

Anion-sensitive regions of L-type CaV1.2 calcium channels expressed in HEK293 cells
- PMID: 20066046
- PMCID: PMC2798859
- DOI: 10.1371/journal.pone.0008602
Anion-sensitive regions of L-type CaV1.2 calcium channels expressed in HEK293 cells
Abstract
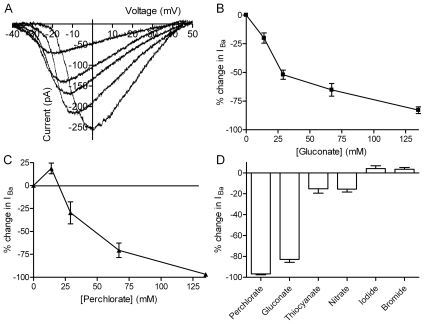
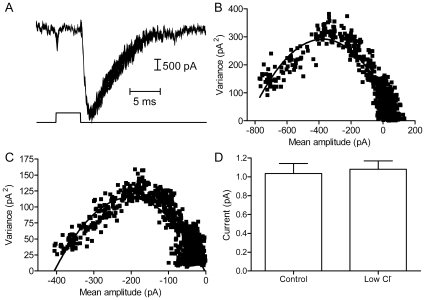
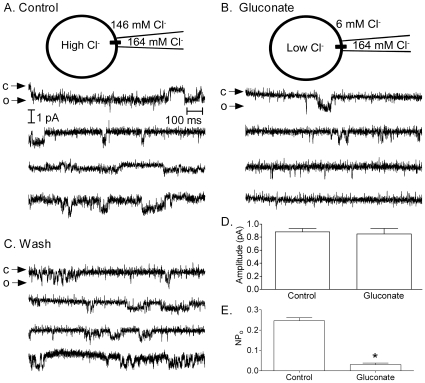
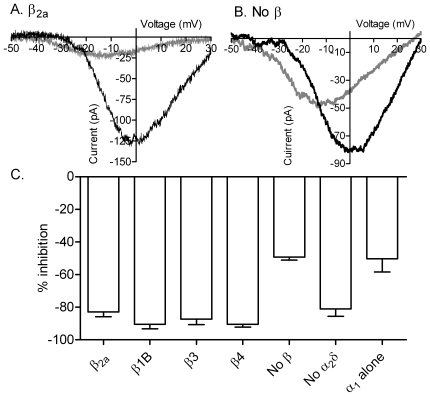
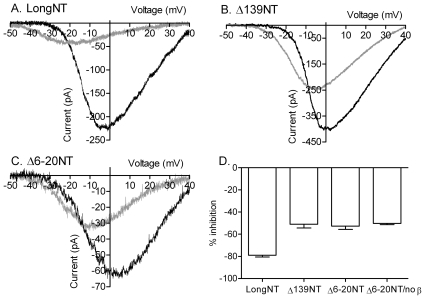
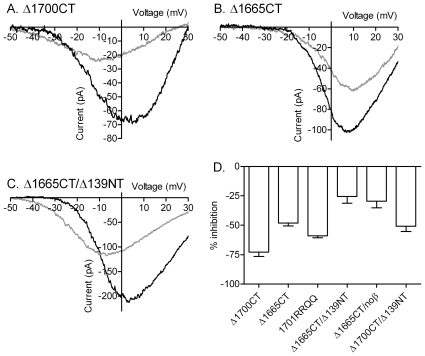
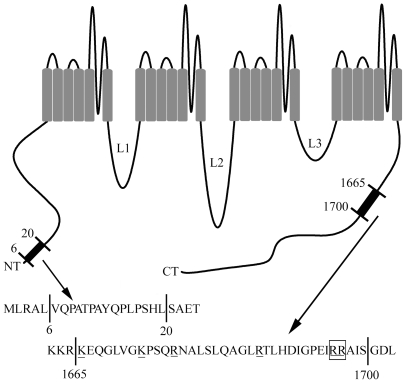
L-type calcium currents (I(Ca)) are influenced by changes in extracellular chloride, but sites of anion effects have not been identified. Our experiments showed that CaV1.2 currents expressed in HEK293 cells are strongly inhibited by replacing extracellular chloride with gluconate or perchlorate. Variance-mean analysis of I(Ca) and cell-attached patch single channel recordings indicate that gluconate-induced inhibition is due to intracellular anion effects on Ca(2+) channel open probability, not conductance. Inhibition of CaV1.2 currents produced by replacing chloride with gluconate was reduced from approximately 75%-80% to approximately 50% by omitting beta subunits but unaffected by omitting alpha(2)delta subunits. Similarly, gluconate inhibition was reduced to approximately 50% by deleting an alpha1 subunit N-terminal region of 15 residues critical for beta subunit interactions regulating open probability. Omitting beta subunits with this mutant alpha1 subunit did not further diminish inhibition. Gluconate inhibition was unchanged with expression of different beta subunits. Truncating the C terminus at AA1665 reduced gluconate inhibition from approximately 75%-80% to approximately 50% whereas truncating it at AA1700 had no effect. Neutralizing arginines at AA1696 and 1697 by replacement with glutamines reduced gluconate inhibition to approximately 60% indicating these residues are particularly important for anion effects. Expressing CaV1.2 channels that lacked both N and C termini reduced gluconate inhibition to approximately 25% consistent with additive interactions between the two tail regions. Our results suggest that modest changes in intracellular anion concentration can produce significant effects on CaV1.2 currents mediated by changes in channel open probability involving beta subunit interactions with the N terminus and a short C terminal region.
Conflict of interest statement
Figures
References
-
- Lipscombe D, Helton TD, Xu W. L-type calcium channels: The low down. J Neurophysiol. 2004;92:2633–2641. - PubMed
-
- Mears D. Regulation of insulin secretion in islets of Langerhans by Ca(2+)channels. J Membr Biol. 2004;200:57–66. - PubMed
-
- Kobayashi T, Yamada Y, Fukao M, Tsutsuura M, Tohse N. Regulation of Cav1.2 current: Interaction with intracellular molecules. J Pharmacol Sci. 2007;103:347–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
