

Remodeling epithelial cell organization: transitions between front-rear and apical-basal polarity
- PMID: 20066074
- PMCID: PMC2742086
- DOI: 10.1101/cshperspect.a000513
Remodeling epithelial cell organization: transitions between front-rear and apical-basal polarity
Abstract
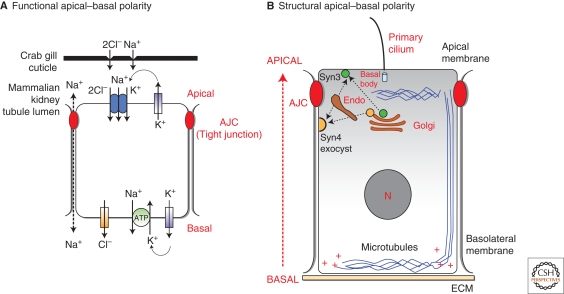
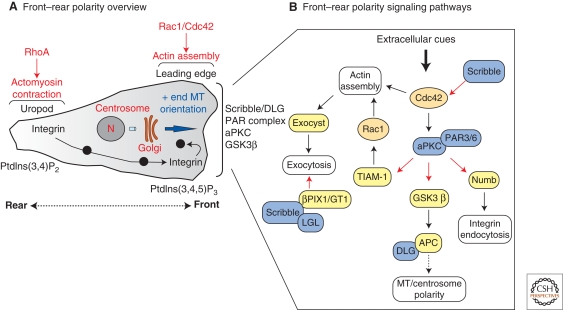
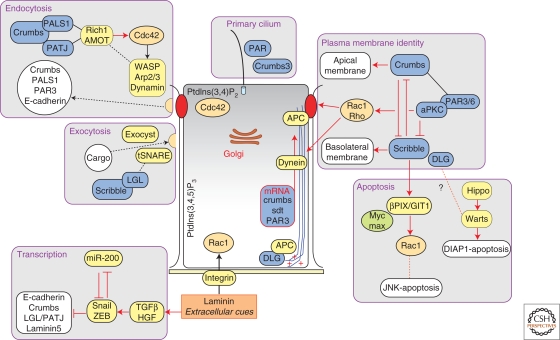
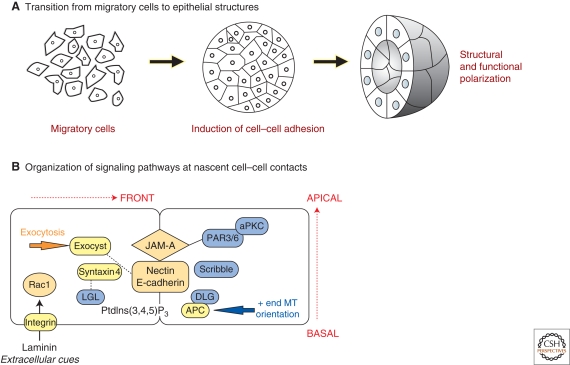
Polarized epithelial cells have a distinctive apical-basal axis of polarity for vectorial transport of ions and solutes across the epithelium. In contrast, migratory mesenchymal cells have a front-rear axis of polarity. During development, mesenchymal cells convert to epithelia by coalescing into aggregates that undergo epithelial differentiation. Signaling networks and protein complexes comprising Rho family GTPases, polarity complexes (Crumbs, PAR, and Scribble), and their downstream effectors, including the cytoskeleton and the endocytic and exocytic vesicle trafficking pathways, together regulate the distributions of plasma membrane and cytoskeletal proteins between front-rear and apical-basal polarity. The challenge is to understand how these regulators and effectors are adapted to regulate symmetry breaking processes that generate cell polarities that are specialized for different cellular activities and functions.
Figures
References
-
- Akhmanova A, Hoogenraad CC, Drabek K, Stepanova T, Dortland B, Verkerk T, Vermeulen W, Burgering BM, De Zeeuw CI, Grosveld F, et al.2001. Clasps are CLIP-115 and -170 associating proteins involved in the regional regulation of microtubule dynamics in motile fibroblasts. Cell 104:923–935 - PubMed
-
- Arimura N, Kaibuchi K 2005. Key regulators in neuronal polarity. Neuron 48:881–884 - PubMed
-
- Audebert S, Navarro C, Nourry C, Chasserot-Golaz S, Lecine P, Bellaiche Y, Dupont JL, Premont RT, Sempere C, Strub JM, et al.2004. Mammalian Scribble forms a tight complex with the betaPIX exchange factor. Curr Biol 14:987–995 - PubMed
-
- Baas AF, Kuipers J, van der Wel NN, Batlle E, Koerten HK, Peters PJ, Clevers HC 2004. Complete polarization of single intestinal epithelial cells upon activation of LKB1 by STRAD. Cell 116:457–466 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources