

The measure of success: constraints, objectives, and tradeoffs in morphogen-mediated patterning
- PMID: 20066078
- PMCID: PMC2742077
- DOI: 10.1101/cshperspect.a002022
The measure of success: constraints, objectives, and tradeoffs in morphogen-mediated patterning
Abstract
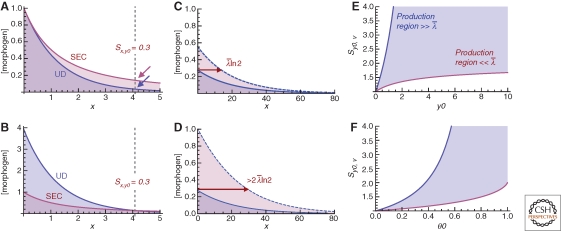
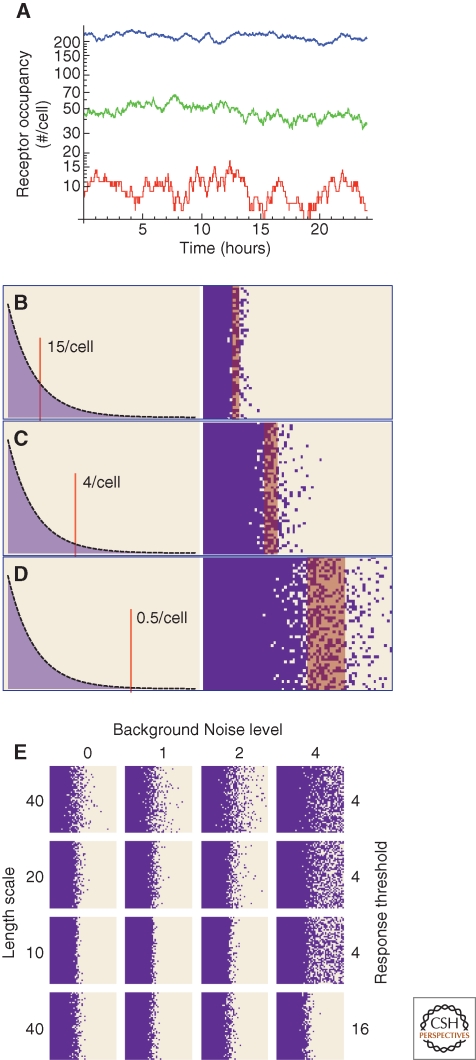
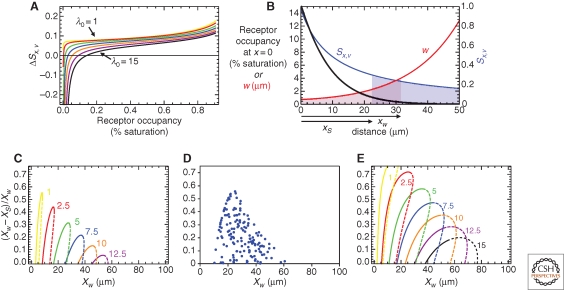
A large, diverse, and growing number of strategies have been proposed to explain how morphogen gradients achieve robustness and precision. We argue that, to be useful, the evaluation of such strategies must take into account the constraints imposed by competing objectives and performance tradeoffs. This point is illustrated through a mathematical and computational analysis of the strategy of self-enhanced morphogen clearance. The results suggest that the usefulness of this strategy comes less from its ability to increase robustness to morphogen source fluctuations per se, than from its ability to overcome specific kinds of noise, and to increase the fraction of a morphogen gradient within which robust threshold positions may be established. This work also provides new insights into the longstanding question of why morphogen gradients show a maximum range in vivo.
Figures
References
-
- Adachi-Yamada T, O'Connor MB 2002. Morphogenetic apoptosis: a mechanism for correcting discontinuities in morphogen gradients. Developmental Biol 251:74–90 - PubMed
-
- Amonlirdviman K, Khare NA, Tree DR, Chen WS, Axelrod JD, Tomlin CJ 2005. Mathematical modeling of planar cell polarity to understand domineering nonautonomy. Science 307:423–426 - PubMed
-
- Ashe HL, Briscoe J 2006. The interpretation of morphogen gradients. Development 133:385–394 - PubMed
-
- Ben-Zvi D, Shilo BZ, Fainsod A, Barkai N 2008. Scaling of the BMP activation gradient in Xenopus embryos. Nature 453:1205–1211 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases