

Gradients in the brain: the control of the development of form and function in the cerebral cortex
- PMID: 20066088
- PMCID: PMC2742095
- DOI: 10.1101/cshperspect.a002519
Gradients in the brain: the control of the development of form and function in the cerebral cortex
Abstract
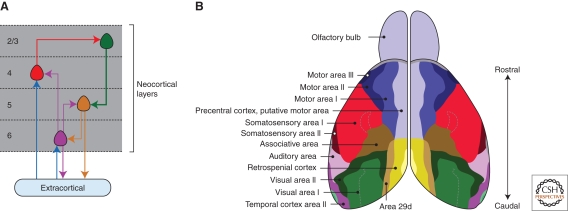
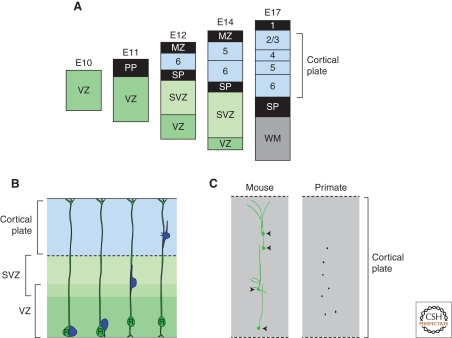
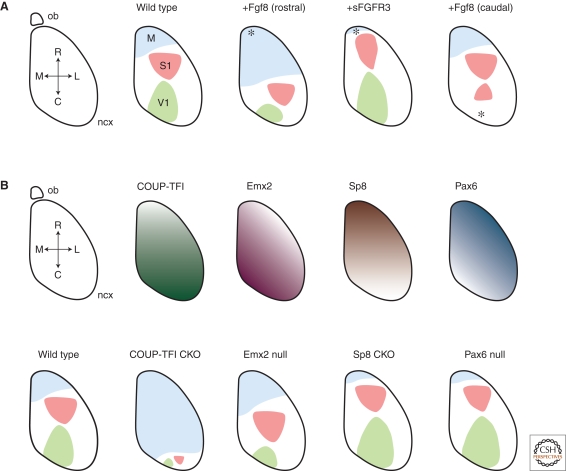
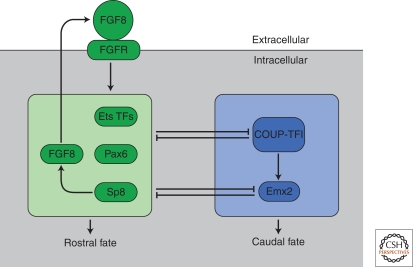
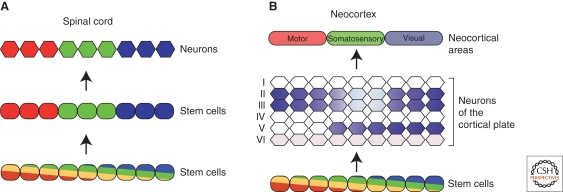
In the developing brain, gradients are commonly used to divide neurogenic regions into distinct functional domains. In this article, we discuss the functions of morphogen and gene expression gradients in the assembly of the nervous system in the context of the development of the cerebral cortex. The cerebral cortex is a mammal-specific region of the forebrain that functions at the top of the neural hierarchy to process and interpret sensory information, plan and organize tasks, and to control motor functions. The mature cerebral cortex is a modular structure, consisting of anatomically and functionally distinct areas. Those areas of neurons are generated from a uniform neuroepithelial sheet by two forms of gradients: graded extracellular signals and a set of transcription factor gradients operating across the field of neocortical stem cells. Fgf signaling from the rostral pole of the cerebral cortex sets up gradients of expression of transcription factors by both activating and repressing gene expression. However, in contrast to the spinal cord and the early Drosophila embryo, these gradients are not subsequently resolved into molecularly distinct domains of gene expression. Instead, graded information in stem cells is translated into discrete, region-specific gene expression in the postmitotic neuronal progeny of the stem cells.
Figures
References
-
- Anderson SA, Eisenstat DD, Shi L, Rubenstein JL 1997. Interneuron migration from basal forebrain to neocortex: Dependence on Dlx genes. Science 278:474–476 - PubMed
-
- Armentano M, Chou SJ, Tomassy GS, Leingartner A, O'Leary DD, Studer M 2007. COUP-TFI regulates the balance of cortical patterning between frontal/motor and sensory areas. Nat Neurosci 10:1277–1286 - PubMed
-
- Bachler M, Neubüser A 2001. Expression of members of the Fgf family and their receptors during midfacial development. Mech Dev 100:313–316 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources