

Review
doi: 10.1101/cshperspect.a002881.
Wnt signaling from development to disease: insights from model systems
Affiliations
- PMID: 20066091
- PMCID: PMC2742092
- DOI: 10.1101/cshperspect.a002881
Item in Clipboard
Review
Wnt signaling from development to disease: insights from model systems
Cold Spring Harb Perspect Biol.
2009 Aug.
Abstract
One of the early surprises in the study of cell adhesion was the discovery that beta-catenin plays dual roles, serving as an essential component of cadherin-based cell-cell adherens junctions and also serving as the key regulated effector of the Wnt signaling pathway. Here, we review our current model of Wnt signaling and discuss how recent work using model organisms has advanced our understanding of the roles Wnt signaling plays in both normal development and in disease. These data help flesh out the mechanisms of signaling from the membrane to the nucleus, revealing new protein players and providing novel information about known components of the pathway.
Figures

A simplified current view of Wnt signaling. See text for details.

Model depicting Wg transcytosis and how this process could influence short- and long-range Wg signaling in the wing imaginal disc. Wg is secreted apically from producing cells, where association with extracellular glypican Dally-like directs it into endocytic vesicles. These Reggie-1-associated vesicles undergo transcytosis and fuse with the plasma membrane to release Wg on the basolateral side of the epithelial sheet. There, it can undergo long-range diffusion and signaling by associating with lipoproteins. See text for further explanation.
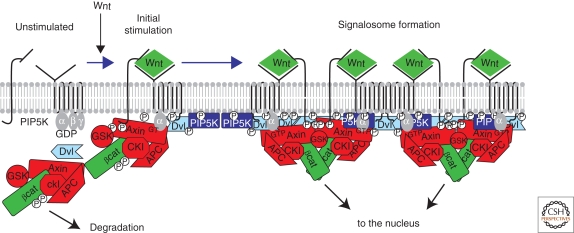
Model for the Wnt-induced formation of LRP5/6 signalosomes. In the absence of Wnt (at the left), LRP and Fz receptors are unassociated in the plasma membrane. Some data suggest that Fz may be coupled to trimeric G proteins, with the Gαo subunit in the inactive GDP-bound form. PIPKIIα (not shown for simplicity) and PIP5KI are membrane associated but inactive. The destruction complex (Axin, APC, GSK, and CKI) phosphorylates βcat, tagging the protein for ubiquitination and proteosomal degradation (see text for further explanation). Initially, Wnt promotes association of LRP5/6 and Fz receptor, leading to phosphorylation of the cytoplasmic tail of LRP5/6 by CKI and GSK3 and recruitment of the destruction complex by binding to Axin. In addition, Gαo is converted to its active GTP-bound form. One potential target of Gαo-GTP is Dvl, which is recruited to the receptor complex, where it can bind and activate PIP5KI, causing an increase in PIP2. All of these modifications appear to reinforce each other in a complex positive feedback loop, leading to a large complex of Wnt, LRP5/6, Fz, Dvl, and degradation complex at the cell surface. GSK activity is inhibited by interaction with the LRP phosphorylated tail, resulting in the accumulation of hypophosphorylated βcat, which can then translocate to the nucleus. See text for more details.

Four proposed models for the role of APC in the destruction complex. (A) APC binds the destruction complex and unknown protein X, recruiting the destruction complex to the apical cell cortex and connecting it to the E3 ubiquitin ligase. (B) The catalytic cycle model in which βcat is sequentially bound to Axin and then APC before transfer to the E3 ligase. (C) Cytoplasmic retention model. Axin and APC bind cytoplasmic βcat and tether it there. (D) APC in nucleus. APC exports βcat from nucleus or acts directly at promotors in transcriptional repression. See text for more details.

Model for target gene regulation by Wg signaling in Drosophila. The top part of the figure outlines some of the mechanisms by which target genes are repressed in the absence of Wg signaling. TCF recruits Gro through direct binding, while CtBP is recruited to WREs in an TCF-independent manner. Both corepressors may act by recruiting HDACs. In addition, the ISWI-ACF1 is recruited to the WRE in a TCF-independent way. Although ISWI remains near the WRE, ACF1 spreads across the loci. Low levels of nuclear Armadillo (Arm) are diverted away from TCF by an APC-CtBP complex or Cby. When high levels of Arm accumulate in the nucleus after Wg signaling, Arm binds to TCF, displacing Gro and recruiting additional coactivators. For simplicity, only two are shown in the bottom part of the figure. The histone acetyltransferase CBP is recruited by the carboxy-terminal Arm transactivation domain and causes a widespread increase in histone acetylation at Wg target loci. ISWI and ACF1 are bound less efficiently to acetylated histone chromatin. The amino-terminal transactivation domain of Arm recruits Lgs and Pygo. Pygo then contacts the Med12 and Med13 subunits of the mediator complex, as well as the TAF4 subunit of the TFIID basal transcription complex. Thus, Pygo acts as an adaptor, connecting the TCF activation complex with the basal transcription machinery to activate target gene expression.
References
-
- Ahmed Y, Hayashi S, Levine A, Wieschaus E 1998. Regulation of Armadillo by a Drosophila APC inhibits neuronal apoptosis during retinal development. Cell 93:1171–1182 - PubMed
-
- Ahmed Y, Nouri A, Wieschaus E 2002. Drosophila Apc1 and Apc2 regulate Wingless transduction throughout development. Development 129:1751–1762 - PubMed
-
- Akong K, Grevengoed E, Price M, McCartney B, Hayden M, DeNofrio J, Peifer M 2002a. Drosophila APC2 and APC1 play overlapping roles in wingless signaling in the embryo and imaginal discs. Dev Biol 250:91–100 - PubMed
-
- Akong K, McCartney B, Peifer M 2002b. Drosophila APC2 and APC1 have overlapping roles in the larval brain despite their distinct intracellular localizations. Dev Biol 250:71–90 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases