

The signaling mechanisms underlying cell polarity and chemotaxis
- PMID: 20066099
- PMCID: PMC2773618
- DOI: 10.1101/cshperspect.a002980
The signaling mechanisms underlying cell polarity and chemotaxis
Abstract
Chemotaxis--the directed movement of cells in a gradient of chemoattractant--is essential for neutrophils to crawl to sites of inflammation and infection and for Dictyostelium discoideum (D. discoideum) to aggregate during morphogenesis. Chemoattractant-induced activation of spatially localized cellular signals causes cells to polarize and move toward the highest concentration of the chemoattractant. Extensive studies have been devoted to achieving a better understanding of the mechanism(s) used by a neutrophil to choose its direction of polarity and to crawl effectively in response to chemoattractant gradients. Recent technological advances are beginning to reveal many fascinating details of the intracellular signaling components that spatially direct the cytoskeleton of neutrophils and D. discoideum and the complementary mechanisms that make the cell's front distinct from its back.
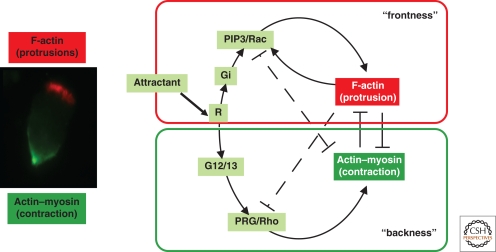
Figures



Similar articles
-
Imaging G-protein coupled receptor (GPCR)-mediated signaling events that control chemotaxis of Dictyostelium discoideum.J Vis Exp. 2011 Sep 20;(55):3128. doi: 10.3791/3128. J Vis Exp. 2011. PMID: 21969095 Free PMC article.
-
Signaling pathways controlling cell polarity and chemotaxis.Trends Biochem Sci. 2001 Sep;26(9):557-66. doi: 10.1016/s0968-0004(01)01934-x. Trends Biochem Sci. 2001. PMID: 11551793 Review.
-
Making all the right moves: chemotaxis in neutrophils and Dictyostelium.Curr Opin Cell Biol. 2004 Feb;16(1):4-13. doi: 10.1016/j.ceb.2003.11.008. Curr Opin Cell Biol. 2004. PMID: 15037299 Review.
-
Teaching resources. Spatial and temporal dynamics of signaling components involved in the control of chemotaxis in Dictyostelium discoideum.Sci STKE. 2004 May 18;2004(234):tr3. doi: 10.1126/stke.2342004tr3. Sci STKE. 2004. PMID: 15161990
-
A G alpha-dependent pathway that antagonizes multiple chemoattractant responses that regulate directional cell movement.Genes Dev. 2004 Apr 1;18(7):805-15. doi: 10.1101/gad.1173404. Epub 2004 Apr 1. Genes Dev. 2004. PMID: 15059962 Free PMC article.
Cited by
-
Tuning Cell Motility via Cell Tension with a Mechanochemical Cell Migration Model.Biophys J. 2020 Jun 16;118(12):2894-2904. doi: 10.1016/j.bpj.2020.04.030. Epub 2020 May 4. Biophys J. 2020. PMID: 32416081 Free PMC article.
-
UNC-6 (netrin) stabilizes oscillatory clustering of the UNC-40 (DCC) receptor to orient polarity.J Cell Biol. 2014 Sep 1;206(5):619-33. doi: 10.1083/jcb.201405026. Epub 2014 Aug 25. J Cell Biol. 2014. PMID: 25154398 Free PMC article.
-
Envisioning migration: mathematics in both experimental analysis and modeling of cell behavior.Curr Opin Cell Biol. 2013 Oct;25(5):538-42. doi: 10.1016/j.ceb.2013.04.004. Epub 2013 May 6. Curr Opin Cell Biol. 2013. PMID: 23660413 Free PMC article. Review.
-
Stimuli-responsive hydrogels for manipulation of cell microenvironment: From chemistry to biofabrication technology.Prog Polym Sci. 2019 Nov;98:101147. doi: 10.1016/j.progpolymsci.2019.101147. Epub 2019 Jul 12. Prog Polym Sci. 2019. PMID: 36467305 Free PMC article.
-
Recent Advances on Cell Culture Platforms for In Vitro Drug Screening and Cell Therapies: From Conventional to Microfluidic Strategies.Adv Healthc Mater. 2023 Jul;12(18):e2202936. doi: 10.1002/adhm.202202936. Epub 2023 Mar 20. Adv Healthc Mater. 2023. PMID: 36898671 Free PMC article. Review.
References
-
- Andrew N, Insall RH 2007. Chemotaxis in shallow gradients is mediated independently of PtdIns 3-kinase by biased choices between random protrusions. Nat Cell Biol 9:193–200 - PubMed
-
- Benard V, Bohl BP, Bokoch GM 1999. Characterization of rac and cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J Biol Chem 274:13198–13204 - PubMed
-
- Bourne HR, Weiner O 2002. A chemical compass. Nature 419:21. - PubMed
-
- Chan TO, Rodeck U, Chan AM, Kimmelman AC, Rittenhouse SE, Panayotou G, Tsichlis PN 2002. Small GTPases and tyrosine kinases coregulate a molecular switch in the phosphoinositide 3-kinase regulatory subunit. Cancer Cell 1:181–191 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources