

Shaping morphogen gradients by proteoglycans
- PMID: 20066107
- PMCID: PMC2773635
- DOI: 10.1101/cshperspect.a002493
Shaping morphogen gradients by proteoglycans
Abstract
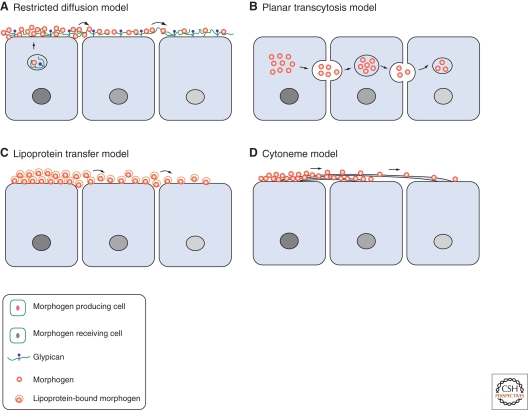
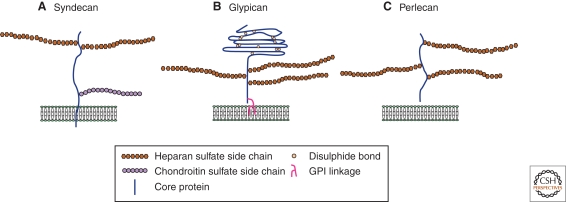
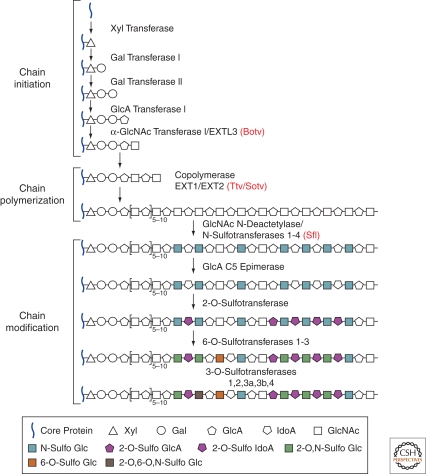
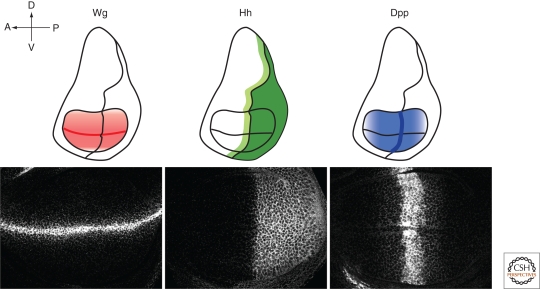
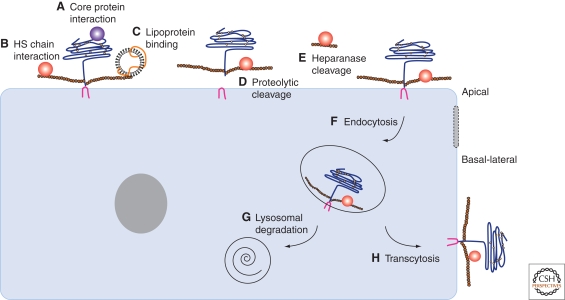
During development, secreted morphogens such as Wnt, Hedgehog (Hh), and BMP emit from their producing cells in a morphogenetic field, and specify different cell fates in a direct concentration-dependent manner. Understanding how morphogens form their concentration gradients to pattern tissues has been a central issue in developmental biology. Various experimental studies from Drosophila have led to several models to explain the formation of morphogen gradients. Over the past decade, one of the main findings in this field is the characterization of heparan sulfate proteoglycan (HSPG) as an essential regulator for morphogen gradient formation. Genetic and cell biological studies have showed that HSPGs can regulate morphogen activities at various steps including control of morphogen movement, signaling, and intracellular trafficking. Here, we review these data, highlighting recent findings that reveal mechanistic roles of HSPGs in controlling morphogen gradient formation.
Figures
References
-
- Affolter M, Basler K 2007. The Decapentaplegic morphogen gradient: From pattern formation to growth regulation. Nat Rev Genet 8:663–674 - PubMed
-
- Ahn J, Ludecke HJ, Lindow S, Horton WA, Lee B, Wagner MJ, Horsthemke B, Wells DE 1995. Cloning of the putative tumour suppressor gene for hereditary multiple exostoses (EXT1). Nat Gen 11:137–143 - PubMed
-
- Baeg GH, Lin X, Khare N, Baumgartner S, Perrimon N 2001. Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless. Development 128:87–94 - PubMed
-
- Baeg GH, Selva EM, Goodman RM, Dasgupta R, Perrimon N 2004. The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors. Dev Biol 276:89–100 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous