

Discovering the molecular components of intercellular junctions--a historical view
- PMID: 20066111
- PMCID: PMC2773636
- DOI: 10.1101/cshperspect.a003061
Discovering the molecular components of intercellular junctions--a historical view
Abstract
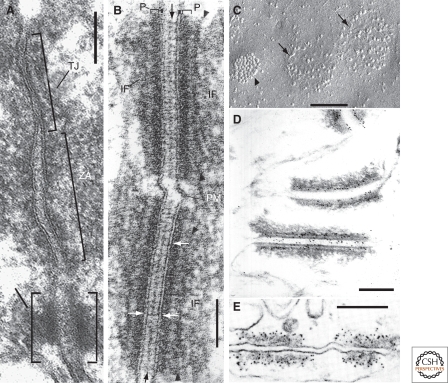
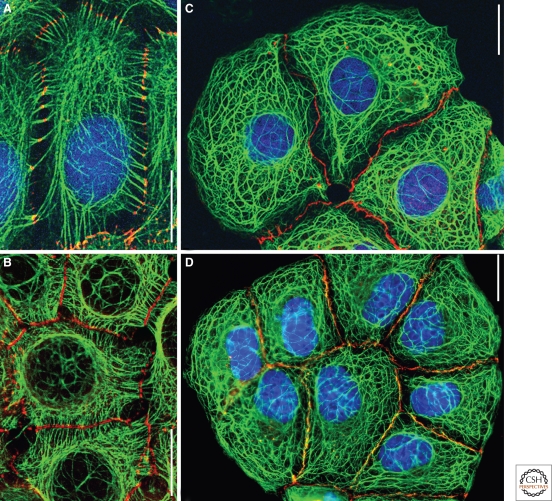
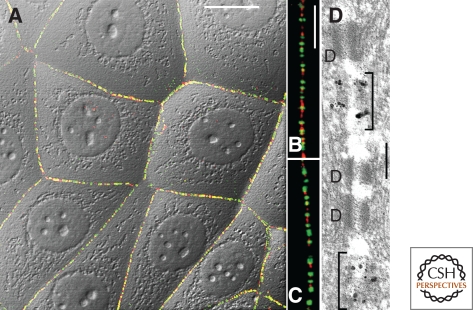
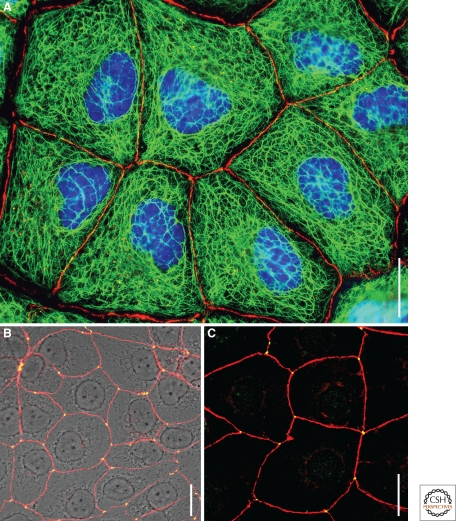
The organization of metazoa is based on the formation of tissues and on tissue-typical functions and these in turn are based on cell-cell connecting structures. In vertebrates, four major forms of cell junctions have been classified and the molecular composition of which has been elucidated in the past three decades: Desmosomes, which connect epithelial and some other cell types, and the almost ubiquitous adherens junctions are based on closely cis-packed glycoproteins, cadherins, which are associated head-to-head with those of the hemi-junction domain of an adjacent cell, whereas their cytoplasmic regions assemble sizable plaques of special proteins anchoring cytoskeletal filaments. In contrast, the tight junctions (TJs) and gap junctions (GJs) are formed by tetraspan proteins (claudins and occludins, or connexins) arranged head-to-head as TJ seal bands or as paracrystalline connexin channels, allowing intercellular exchange of small molecules. The by and large parallel discoveries of the junction protein families are reported.
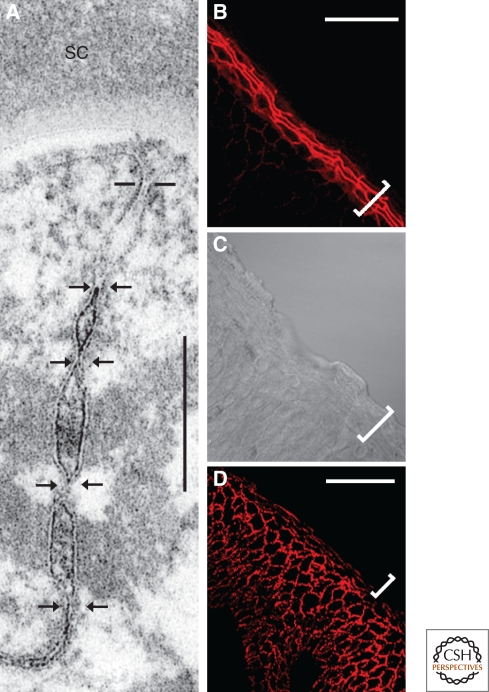
Figures
Similar articles
-
Trans-Compartmental Regulation of Tight Junction Barrier Function.Tissue Barriers. 2023 Oct 2;11(4):2133880. doi: 10.1080/21688370.2022.2133880. Epub 2022 Oct 11. Tissue Barriers. 2023. PMID: 36220768 Free PMC article. Review.
-
Targeting of p0071 to desmosomes and adherens junctions is mediated by different protein domains.J Cell Sci. 2003 Apr 1;116(Pt 7):1219-33. doi: 10.1242/jcs.00275. J Cell Sci. 2003. PMID: 12615965
-
Prostaglandin-induced cervical remodelling in humans in the first trimester is associated with increased expression of specific tight junction, but not gap junction proteins.J Transl Med. 2012 Mar 7;10:40. doi: 10.1186/1479-5876-10-40. J Transl Med. 2012. PMID: 22397627 Free PMC article. Clinical Trial.
-
Connexins, E-cadherin, Claudin-7 and β-catenin transiently form junctional nexuses during the post-natal mammary gland development.Dev Biol. 2016 Aug 1;416(1):52-68. doi: 10.1016/j.ydbio.2016.06.011. Epub 2016 Jun 9. Dev Biol. 2016. PMID: 27291930
-
Gap junctions and connexin-interacting proteins.Cardiovasc Res. 2004 May 1;62(2):233-45. doi: 10.1016/j.cardiores.2003.12.009. Cardiovasc Res. 2004. PMID: 15094344 Review.
Cited by
-
The urothelium: a multi-faceted barrier against a harsh environment.Mucosal Immunol. 2022 Jun;15(6):1127-1142. doi: 10.1038/s41385-022-00565-0. Epub 2022 Sep 30. Mucosal Immunol. 2022. PMID: 36180582 Free PMC article. Review.
-
Transition of podosomes into zipper-like structures in macrophage-derived multinucleated giant cells.Mol Biol Cell. 2020 Aug 15;31(18):2002-2020. doi: 10.1091/mbc.E19-12-0707. Epub 2020 Jun 24. Mol Biol Cell. 2020. PMID: 32579434 Free PMC article.
-
Structure, function, and regulation of desmosomes.Prog Mol Biol Transl Sci. 2013;116:95-118. doi: 10.1016/B978-0-12-394311-8.00005-4. Prog Mol Biol Transl Sci. 2013. PMID: 23481192 Free PMC article. Review.
-
Evolution: functional evolution of nuclear structure.J Cell Biol. 2011 Oct 17;195(2):171-81. doi: 10.1083/jcb.201103171. J Cell Biol. 2011. PMID: 22006947 Free PMC article. Review.
-
Novel mechanism of cytokine-induced disruption of epithelial barriers: Janus kinase and protein kinase D-dependent downregulation of junction protein expression.Tissue Barriers. 2013 Oct 1;1(4):e25231. doi: 10.4161/tisb.25231. Epub 2013 Jun 3. Tissue Barriers. 2013. PMID: 24665409 Free PMC article.
References
-
- Aberle H, Schwartz H, Kemler R 1996. Cadherin-catenin complex: Protein interactions and their implications for cadherin function. J Cell Biochem 61:514–523 - PubMed
-
- Aghib DF, McCrea PD 1995. The E-cadherin complex contains the src substrate p120. Exp Cell Res 218:359–369 - PubMed
-
- Aho S, Rothenberger K, Uitto J 1999. Human p120ctn catenin: Tissue-specific expression of isoforms and molecular interactions with BP180/type XVII collagen. J Cell Biochem 73:390–399 - PubMed
-
- Amagai M 1999. Autoimmunity against desmosomal cadherins in pemphigus. J Dermatol Sci 20:92–102 - PubMed
-
- Amagai M, Klaus-Kovtun V, Stanley JR 1991. Autoantibodies against a novel epithelial cadherin in pemphigus vulgaris, a disease of cell adhesion. Cell 67:869–977 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous