

Review
doi: 10.1101/cshperspect.a000489.
Gradients and the specification of planar polarity in the insect cuticle
Affiliations
- PMID: 20066114
- PMCID: PMC2773641
- DOI: 10.1101/cshperspect.a000489
Item in Clipboard
Review
Gradients and the specification of planar polarity in the insect cuticle
Cold Spring Harb Perspect Biol.
2009 Nov.
Abstract
In addition to specifying cell fate, there is a wealth of evidence that molecular gradients are also primarily responsible for specifying cell polarity, particularly in the plane of epithelial sheets ("planar polarity"). The first compelling evidence of a role for gradients in specifying planar polarity came from transplantation experiments in the insect cuticle. More recent molecular genetic analyses in the fruit fly Drosophila have begun to give insights into the molecular nature of the gradients involved, and how they are interpreted at the cellular level.
Figures

Transplantation experiments and planar polarity in the insect cuticle. (A) Diagram showing the polarity changes observed in the cuticle of the moth Galleria following 180° rotation of a piece of cuticle. Arrows indicate normal direction of polarization of scales from anterior to posterior. The gray box indicates approximate extent of the region of the cuticle that was rotated. (Based on Piepho 1955). (B, D) Diagrams showing the effects on the cuticle ridge pattern on Rhodnius, produced by rotating a piece of cuticle 180° (B) or reciprocally transplanting pieces of cuticle to different positions in the AP axis of a segment (D). Locke proposed that cells at different positions in the AP axis attempt to join up with cells of their original “level” after transplantation (Locke 1959). (C, E) Models based on the concept of a gradient of a diffusible substance specifying planar polarity (Lawrence 1966; Stumpf 1966) to explain the transplantation results of Locke. After transplantation, the gradient profile is altered. Diffusion then modifies the shape of the gradient to provide smooth peaks and troughs. Cells take on their polarity according to the direction of the slope of the gradient (arrows), with the experimental manipulations resulting in reversal of the slope and hence of polarity (dark gray arrows). (F, G) Models for how local cell alignment and the local organizing properties of segment boundaries might specify planar polarity across an insect segment. Cells within the segment (light gray) align their polarities with their immediate neighbors. Cells at the segment boundary (dark gray) might directly polarize the first row of cells in the segment (white arrows, F), with this polarity being passed on cell by cell. Alternatively, cells at the segment boundary might generate a local gradient that polarizes the first few rows of cells, with polarity in the rest of the segment being again determined by local cell–cell interactions (G).

Frizzled and the determination of planar polarity in Drosophila. (A) Diagram showing the effects of loss of fz activity from a clone of cells in the Drosophila wing. Normal hair polarity (arrows) is from proximal to distal; however, around a fz clone (gray), hairs point towards the clone. (B) Groups of photoreceptors (ommatidia, black) show a distinct polarity on the dorsoventral (DV) axis of the eye. A clone of cells lacking fz activity (gray region) in the dorsal half of the eye cause ommatidial dorsal to the clone (i.e., on the “polar” side relative to the dorsoventral midline) to show inverted DV polarity (gray). (C) Model for Fz function in nonautonomously specifying planar polarity, through production of a gradient of a secondary signal. The Fz receptor is activated in a gradient across the axis of the wing by an extracellular ligand gradient (triangles). Fz activity then induces a gradient of a secondary signal which acts nonautonomously (e.g., via diffusion) to specify cell polarity. (D) Model for Fz function through passing a polarized signal from cell to cell. Each cell is proposed to transduce a polarity signal (triangles) via the Fz receptor and respond by polarizing itself and secreting the same signal at the opposite cell edge, which in turn polarizes the neighboring cell. (E) Diagram showing the asymmetric localization of Fz and the core polarity proteins in the junctional regions of pupal wing cells. Fz and a number of other proteins localize in a junctional complex at the distal cell edge (light gray), whereas a reciprocal complex of different composition forms at the proximal cell edge (dark gray). (F) Asymmetric localization of Fz and the core polarity proteins in the R3/R4 photoreceptor pair and the determination of ommatidial polarity. Early in ommatidial assembly, the ommatidium consists of five photoreceptors (R3, R4, R5, R8, and R2) and is polarized only on the AP axis. Within the most anterior pair of photoreceptors (R3/R4), Fz becomes localized to the R3/R4 cell–cell boundary on the side closest to the equatorial midline of the eye (dotted line), causing the cell closest to the midline to take on R3 fate and the cell further away to take on R4 fate. As a result of this cell fate decision, ommatidia dorsal to the midline rotate clockwise and those ventral rotate anticlockwise, resulting in ommatidia with opposite DV polarities. (G) Model to explain nonautonomous polarity changes around an fz clone in the wing, based on coupling of cell polarities via junctional assembly of asymmetric core planar polarity complexes. Cells within the clone cannot form Fz-containing complexes, and so form the reciprocal proximal complex (dark gray) with cells surrounding the clone. These cells in turn assemble Fz-containing distal complexes (light gray) around the clone edges, and this abnormal polarity of asymmetric complex assembly is propagated to surround cells, such that they take on abnormal polarity.
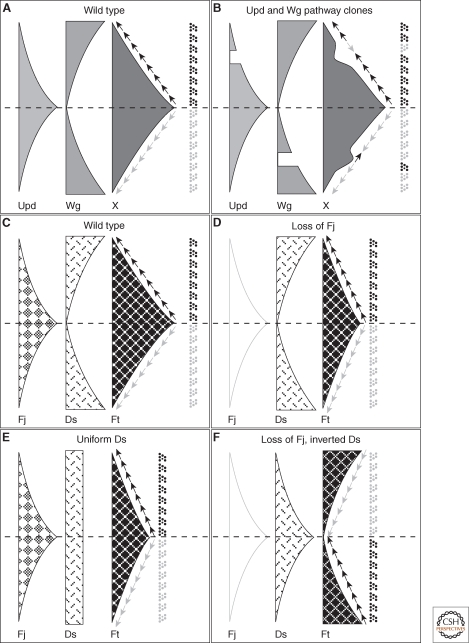
Gradients and the specification of ommatidial DV polarity in the Drosophila eye. (A) Model for the activities of the Unpaired (Upd) and Wg pathways in the eye, regulating a graded factor ("X") that specifies ommatidial polarity. The Upd ligand is expressed at the equator and forms a gradient of activation of the JAK/STAT signal transduction pathway (light gray), whereas Wg is expressed at the dorsal and ventral poles of the eye and similarly generates a gradient of pathway activation (mid gray). A nonautonomously acting factor X (dark gray) is positively regulated by Upd signaling and negatively regulated by Wg signaling. The slope of X specifies ommatidial DV polarity (arrows). (B) Model to explain the phenotypes of clones that block Upd or Wg signal transduction. A clone blocking Upd transduction (upper part of panel) causes a trough in the gradient of X, causing a change in the slope on the polar side of the clone and polar inversions of ommatidial polarity. A clone blocking Wg transduction (lower part of panel) causes a peak in the gradient of X, and inversion of ommatidial polarity on the equatorial side of the clone. (C) Model showing equatorial-to-polar gradient of Four-jointed (Fj) expression and polar-to-equatorial gradient of Dachsous (Ds) expression acting together to produce an equatorial-to-polar gradient of Fat (Ft) activity (Ft being uniformly expressed). Ommatidial polarity is specified by the slope of the gradient of Ft activity (arrows). (D) Loss of Fj does not cause significant ommatidial polarity defects, as the Ds expression gradient is sufficient to provide graded Ft activity. (E) Uniform expression of Ds, with graded Fj expression, is also sufficient to provide graded Ft activity. However, Ds expression cannot be dispensed with, as Fj acts by modulating the Ft/Ds interaction and in the absence of Ds, Fj cannot modulate Ft activity. (F) Loss of Fj and inversion of the Ds gradient causes inversion of ommatidial polarities.

Gradients in the Drosophila abdomen. The abdomen is composed of a series of repeating segments, from anterior to posterior, each divided into anterior (A) and posterior (P) compartments at the A/P compartment boundary (dotted line). Adjacent segments are separated by the segmental boundary that also corresponds to the P/A compartment boundary. The photograph (top) shows a single segment, anterior to the left, posterior to the right, with hairs and bristles pointing posteriorly. Diagrams show putative gradients of factors involved in patterning fate and polarity in the abdomen, with the anterior compartment on the left and the posterior compartment on the right.

Models of gradient function and the polarization of individual cells. (A) Models for graded Fz activity being converted into Fz subcellular asymmetry via formation of asymmetric junctional complexes. An extrinsic gradient is assumed to produce a gradient of activated Fz across the tissue. For any particular cell, the next more proximal cell (left) has a higher concentration of activated Fz than the next more distal cell (right); hence, “proximal” junctional complex components (dark gray) in this cell prefer to form asymmetric complexes with “distal” Fz-containing complexes (light gray) in the more proximal cell. Consequently, each cell becomes polarized with proximal and distal complexes at opposite ends. If only “activated” Fz can participate in complex formation, then a gradient of amounts of protein in proximal and distal complexes would be expected (middle row of cells in diagram), but this has not been experimentally observed. If total cellular Fz can incorporate into complexes, once asymmetry has been established, then all cells will show the same concentration of complexes at junctions (bottom row of cells in diagram), which more closely fits the experimental observations. (B) Model in which cells compare levels of Fz activity, before establishment of Fz asymmetric localization, showing the effects of a clone of fz mutant cells. An Fz activity gradient is established in the tissue, after which cells compare their levels with their neighbors and take on an average value. As fz mutant cells have no Fz activity, this leads neighboring cells to take on a lower than normal level of Fz activity, causing an inversion of the gradient of Fz activity distally (to the right) of the clone. The different levels of Fz activity between neighboring cells then drive asymmetric subcellular localization of Fz via an unknown mechanism, possibly similar to that in panel (A). (C) Model for how Ds and Fj gradients might polarize cells. A gradient of Ds expression leads to a gradient of formation of Ds-Ft heterodimers between the edges of each cell. An opposing gradient of Fj expression (not shown) promotes this process by modulating the strength of Ds-Ft binding. By measuring the ratio of heterodimers between each cell edge, each cell can obtain a polarity cue. This cue would be expected to be an initially subtle gradient of heterodimer formation (third row in diagram), but might ultimately be amplified to provide a strong polarity cue (bottom row in diagram). (D) Application of the Ds-Ft heterodimer model to explain the nonautonomy seen around ds clones. Cells lacking ds can only form Ft-Ds heterodimers and not Ds-Ft heterodimers with their neighbors, which affects the availability of Ds in the neighboring cells for forming heterodimers with other neighbors. If cells determine their polarity by measuring the ratio of Ft-Ds to Ds-Ft heterodimers on their cell edges, then this would cause an inversion of cell polarity on the right side of the clone.
Similar articles
-
Cell interactions and planar polarity in the abdominal epidermis of Drosophila.Development. 2004 Oct;131(19):4651-64. doi: 10.1242/dev.01351. Epub 2004 Aug 25. Development. 2004. PMID: 15329345
-
Asymmetric localization of frizzled and the establishment of cell polarity in the Drosophila wing.Mol Cell. 2001 Feb;7(2):367-75. doi: 10.1016/s1097-2765(01)00184-8. Mol Cell. 2001. PMID: 11239465
-
Planar cell polarity in the Drosophila eye is directed by graded Four-jointed and Dachsous expression.Development. 2004 Dec;131(24):6175-84. doi: 10.1242/dev.01550. Epub 2004 Nov 17. Development. 2004. PMID: 15548581
-
Frizzled signaling and the developmental control of cell polarity.Trends Genet. 1998 Nov;14(11):452-8. doi: 10.1016/s0168-9525(98)01584-4. Trends Genet. 1998. PMID: 9825673 Review.
-
The frizzled/stan pathway and planar cell polarity in the Drosophila wing.Curr Top Dev Biol. 2012;101:1-31. doi: 10.1016/B978-0-12-394592-1.00001-6. Curr Top Dev Biol. 2012. PMID: 23140623 Free PMC article. Review.
Cited by
-
Planar cell polarity: global inputs establishing cellular asymmetry.Curr Opin Cell Biol. 2017 Feb;44:110-116. doi: 10.1016/j.ceb.2016.08.002. Epub 2016 Aug 26. Curr Opin Cell Biol. 2017. PMID: 27576155 Free PMC article. Review.
-
Wnt/PCP proteins regulate stereotyped axon branch extension in Drosophila.Development. 2012 Jan;139(1):165-77. doi: 10.1242/dev.068668. Development. 2012. PMID: 22147954 Free PMC article.
-
Planar cell polarity signaling in the Drosophila eye.Curr Top Dev Biol. 2010;93:189-227. doi: 10.1016/B978-0-12-385044-7.00007-2. Curr Top Dev Biol. 2010. PMID: 20959167 Free PMC article. Review.
-
A theoretical framework for planar polarity establishment through interpretation of graded cues by molecular bridges.Development. 2019 Feb 1;146(3):dev168955. doi: 10.1242/dev.168955. Development. 2019. PMID: 30709912 Free PMC article. Review.
-
An extended steepness model for leg-size determination based on Dachsous/Fat trans-dimer system.Sci Rep. 2014 Mar 11;4:4335. doi: 10.1038/srep04335. Sci Rep. 2014. PMID: 24613915 Free PMC article.
References
-
- Adler PN, Krasnow RE, Liu J 1997. Tissue polarity points from cells that have higher Frizzled levels towards cells that have lower Frizzled levels. Curr Biol 7:940–949 - PubMed
-
- Adler P, Charlton J, Liu J 1998. Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development 125:959–968 - PubMed
-
- Amonlirdviman K, Khare NA, Tree DRP, Chen W-S, Axelrod JD, Tomlin CJ 2005. Mathematical modeling of planar cell polarity to understand domineering non-autonomy. Science 307:423–426 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases