

Symmetry breaking in plants: molecular mechanisms regulating asymmetric cell divisions in Arabidopsis
- PMID: 20066115
- PMCID: PMC2773642
- DOI: 10.1101/cshperspect.a000497
Symmetry breaking in plants: molecular mechanisms regulating asymmetric cell divisions in Arabidopsis
Abstract
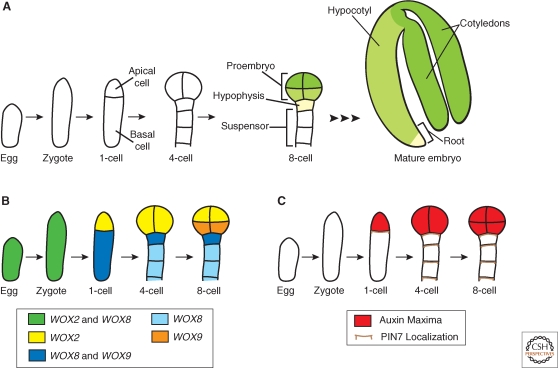
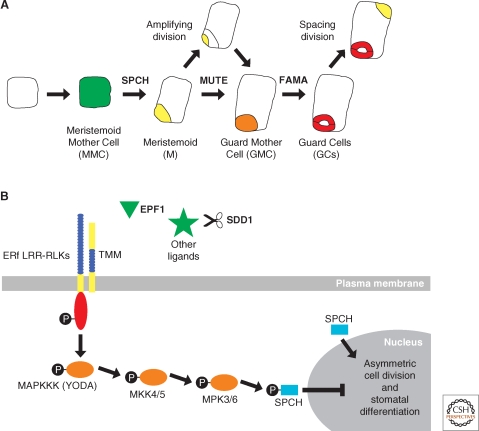
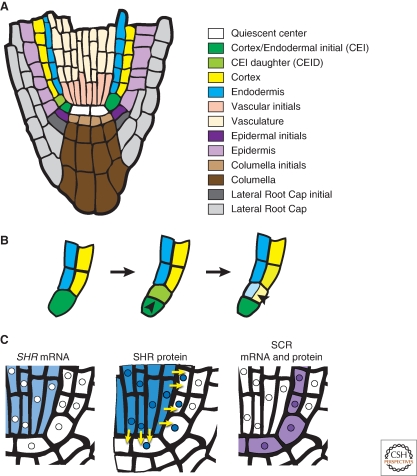
Asymmetric cell division generates cell types with different specialized functions or fates. This type of division is critical to the overall cellular organization and development of many multicellular organisms. In plants, regulated asymmetric cell divisions are of particular importance because cell migration does not occur. The influence of extrinsic cues on asymmetric cell division in plants is well documented. Recently, candidate intrinsic factors have been identified and links between intrinsic and extrinsic components are beginning to be elucidated. A novel mechanism in breaking symmetry was revealed that involves the movement of typically intrinsic factors between plant cells. As we learn more about the regulation of asymmetric cell divisions in plants, we can begin to reflect on the similarities and differences between the strategies used by plants and animals. Focusing on the underlying molecular mechanisms, this article describes three selected cases of symmetry-breaking events in the model plant Arabidopsis thaliana. These examples occur in early embryogenesis, stomatal development, and ground tissue formation in the root.
Figures
References
-
- Alessa L, Kropf DL 1999. F-actin marks the rhizoid pole in living Pelvetia compressa zygotes. Development 126:201–209 - PubMed
-
- Balasubramanian MK, Bi E, Glotzer M 2004. Comparative analysis of cytokinesis in budding yeast, fission yeast and animal cells. Curr Biol 14:R806–818 - PubMed
-
- Benfey PN, Linstead PJ, Roberts K, Schiefelbein JW, Hauser MT, Aeschbacher RA 1993. Root development in Arabidopsis: Four mutants with dramatically altered root morphogenesis. Development 119:57–70 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources