

Glycosidase inhibition: assessing mimicry of the transition state
- PMID: 20066263
- PMCID: PMC2822703
- DOI: 10.1039/b915870g
Glycosidase inhibition: assessing mimicry of the transition state
Abstract
Glycoside hydrolases, the enzymes responsible for hydrolysis of the glycosidic bond in di-, oligo- and polysaccharides, and glycoconjugates, are ubiquitous in Nature and fundamental to existence. The extreme stability of the glycosidic bond has meant these enzymes have evolved into highly proficient catalysts, with an estimated 10(17) fold rate enhancement over the uncatalysed reaction. Such rate enhancements mean that enzymes bind the substrate at the transition state with extraordinary affinity; the dissociation constant for the transition state is predicted to be 10(-22) M. Inhibition of glycoside hydrolases has widespread application in the treatment of viral infections, such as influenza and HIV, lysosomal storage disorders, cancer and diabetes. If inhibitors are designed to mimic the transition state, it should be possible to harness some of the transition state affinity, resulting in highly potent and specific drugs. Here we examine a number of glycosidase inhibitors which have been developed over the past half century, either by Nature or synthetically by man. A number of criteria have been proposed to ascertain which of these inhibitors are true transition state mimics, but these features have only be critically investigated in a very few cases.
Figures


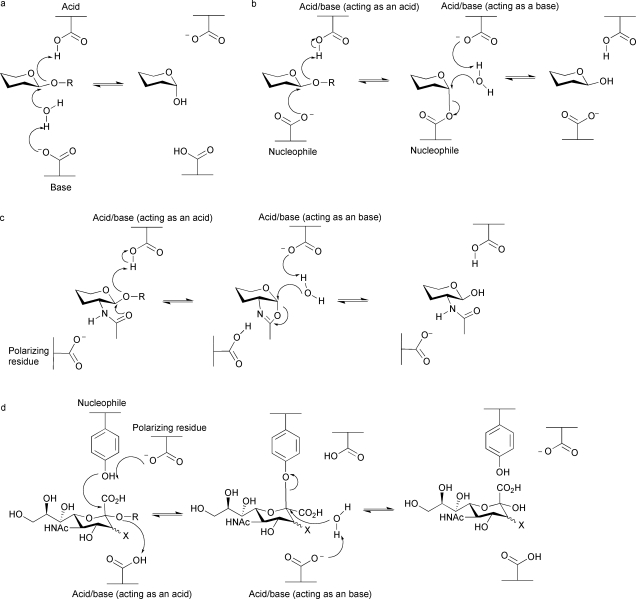
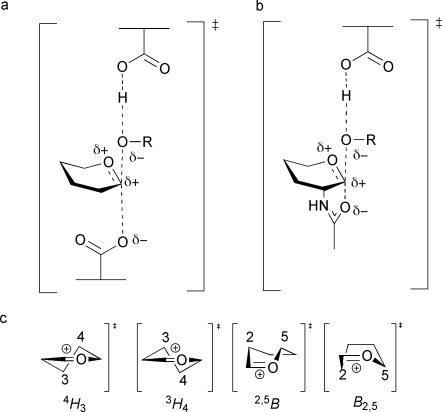
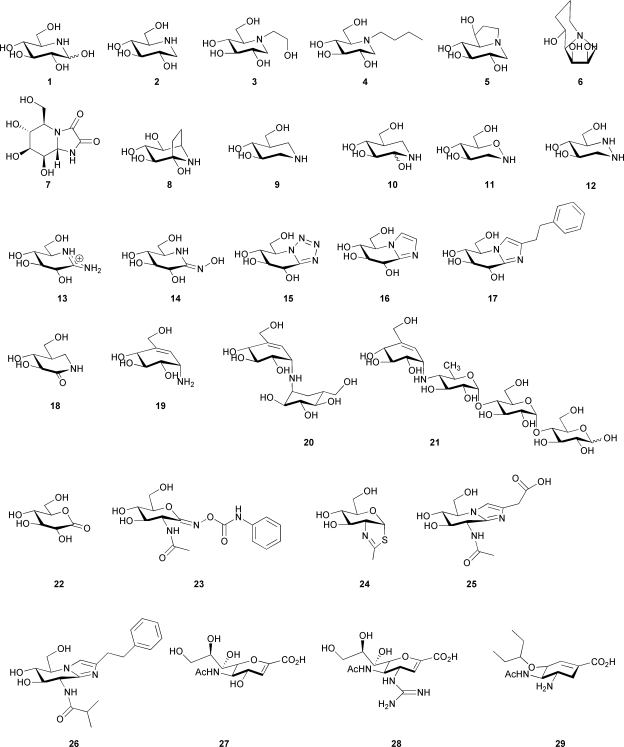
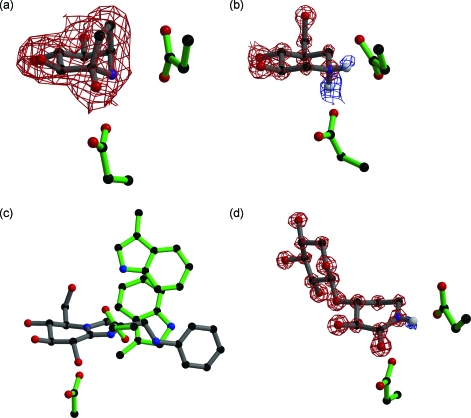
References
-
- Davies G. J., Gloster T. M., Henrissat B. Curr. Opin. Struct. Biol. 2005;15:637–645. - PubMed
-
- Laine R. A. Glycobiology. 1994;4:759–767. - PubMed
-
- Wolfenden R., Lu X., Young G. J. Am. Chem. Soc. 1998;120:6814–6815.
-
- Suzuki Y., Ichinomiya S., Kurosawa M., Ohkubo M., Watanabe H., Iwasaki H., Matsuda J., Noguchi Y., Takimoto K., Itoh M., Tabe M., Iida M., Kubo T., Ogawa S., Nanbe E., Higaki K., Ohno K., Brady R. O. Ann. Neurol. 2007;62:671–675. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous