

Priming of immune responses against transporter associated with antigen processing (TAP)-deficient tumours: tumour direct priming
- PMID: 20067541
- PMCID: PMC2770689
- DOI: 10.1111/j.1365-2567.2009.03127.x
Priming of immune responses against transporter associated with antigen processing (TAP)-deficient tumours: tumour direct priming
Abstract
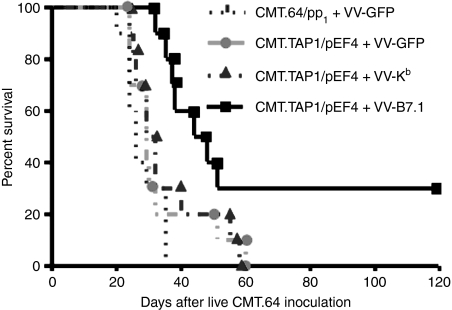
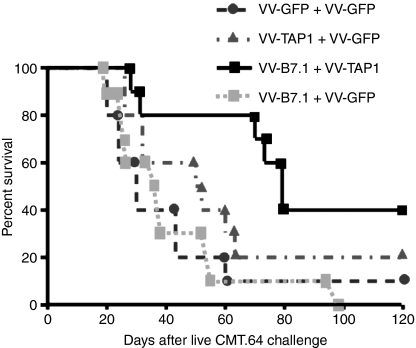
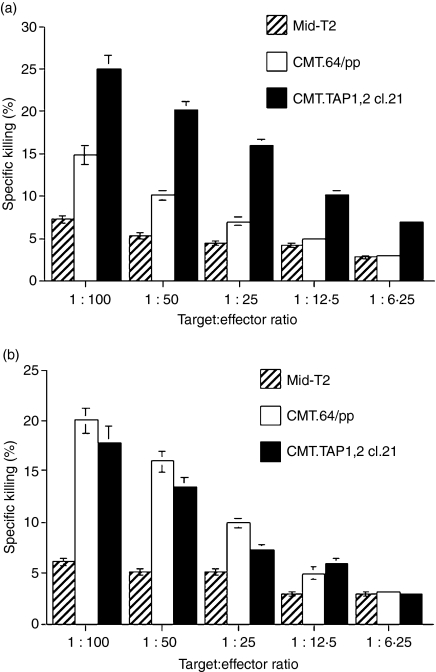
We previously showed that introduction of transporter associated with antigen processing (TAP) 1 into TAP-negative CMT.64, a major histocompatibility complex class I (MHC-I) down-regulated mouse lung carcinoma cell line, enhanced T-cell immunity against TAP-deficient tumour cells. Here, we have addressed two questions: (1) whether such immunity can be further augmented by co-expression of TAP1 with B7.1 or H-2K(b) genes, and (2) which T-cell priming mechanism (tumour direct priming or dendritic cell cross-priming) plays the major role in inducing an immune response against TAP-deficient tumours. We introduced the B7.1 or H-2K(b) gene into TAP1-expressing CMT.64 cells and determined which gene co-expressed with TAP1 was able to provide greater protective immunity against TAP-deficient tumour cells. Our results show that immunization of mice with B7.1 and TAP1 co-expressing but not H-2K(b) and TAP1 co-expressing CMT.64 cells dramatically augments T-cell-mediated immunity, as shown by an increase in survival of mice inoculated with live CMT.64 cells. In addition, our results suggest that induction of T-cell-mediated immunity against TAP-deficient tumour cells could be mainly through tumour direct priming rather than dendritic cell cross-priming as they show that T cells generated by tumour cell-lysate-loaded dendritic cells recognized TAP-deficient tumour cells much less than TAP-proficient tumour cells. These data suggest that direct priming by TAP1 and B7.1 co-expressing tumour cells is potentially a major mechanism to facilitate immune responses against TAP-deficient tumour cells.
Figures

Similar articles
-
Effect of B7.1 costimulation on T-cell based immunity against TAP-negative cancer can be facilitated by TAP1 expression.PLoS One. 2009 Jul 24;4(7):e6385. doi: 10.1371/journal.pone.0006385. PLoS One. 2009. PMID: 19629186 Free PMC article.
-
Activation of antigen-specific cytotoxic T lymphocytes by beta 2-microglobulin or TAP1 gene disruption and the introduction of recipient-matched MHC class I gene in allogeneic embryonic stem cell-derived dendritic cells.J Immunol. 2008 Nov 1;181(9):6635-43. doi: 10.4049/jimmunol.181.9.6635. J Immunol. 2008. PMID: 18941254
-
GM-CSF and B7-1 (CD80) co-stimulatory signals co-operate in the induction of effective anti-tumor immunity in syngeneic mice.Int J Cancer. 1997 Nov 14;73(4):556-61. doi: 10.1002/(sici)1097-0215(19971114)73:4<556::aid-ijc17>3.0.co;2-7. Int J Cancer. 1997. PMID: 9389572
-
Enhancement of effector CD8+ T-cell function by tumour-associated B7-H3 and modulation of its counter-receptor triggering receptor expressed on myeloid cell-like transcript 2 at tumour sites.Immunology. 2010 Jul;130(3):363-73. doi: 10.1111/j.1365-2567.2009.03236.x. Epub 2010 Feb 5. Immunology. 2010. PMID: 20141543 Free PMC article.
-
Adaptive immunity in cancer immunology and therapeutics.Ecancermedicalscience. 2014 Jul 2;8:441. doi: 10.3332/ecancer.2014.441. eCollection 2014. Ecancermedicalscience. 2014. PMID: 25075215 Free PMC article. Review.
Cited by
-
A novel category of antigens enabling CTL immunity to tumor escape variants: Cinderella antigens.Cancer Immunol Immunother. 2012 Jan;61(1):119-25. doi: 10.1007/s00262-011-1160-x. Epub 2011 Nov 25. Cancer Immunol Immunother. 2012. PMID: 22116347 Free PMC article. Review.
-
[Paclitaxel blocks immunologic escape through up-regulating TAP-1, TAP-2 and eliminatiing Treg cells in 3LL-bearing mice].Zhongguo Fei Ai Za Zhi. 2010 Oct;13(10):937-41. doi: 10.3779/j.issn.1009-3419.2010.10.13. Zhongguo Fei Ai Za Zhi. 2010. PMID: 20959065 Free PMC article. Chinese.
-
Limited density of an antigen presented by RMA-S cells requires B7-1/CD28 signaling to enhance T-cell immunity at the effector phase.PLoS One. 2014 Nov 10;9(11):e108192. doi: 10.1371/journal.pone.0108192. eCollection 2014. PLoS One. 2014. PMID: 25383875 Free PMC article.
-
Dendritic cell based PSMA immunotherapy for prostate cancer using a CD40-targeted adenovirus vector.PLoS One. 2012;7(10):e46981. doi: 10.1371/journal.pone.0046981. Epub 2012 Oct 8. PLoS One. 2012. PMID: 23056548 Free PMC article.
-
Downregulation of expression of transporters associated with antigen processing 1 and 2 and human leukocyte antigen I and its effect on immunity in nasopharyngeal carcinoma patients.Mol Clin Oncol. 2014 Jan;2(1):51-58. doi: 10.3892/mco.2013.194. Epub 2013 Sep 26. Mol Clin Oncol. 2014. PMID: 24649307 Free PMC article.
References
-
- Abele R, Tampe R. The ABCs of immunology: structure and function of TAP, the transporter associated with antigen processing. Physiology (Bethesda) 2004;19:216–24. - PubMed
-
- Seliger B, Maeurer MJ, Ferrone S. TAP off – tumors on. Immunol Today. 1997;18:292–9. - PubMed
-
- Seliger B, Maeurer MJ, Ferrone S. Antigen-processing machinery breakdown and tumor growth. Immunol Today. 2000;21:455–64. - PubMed
-
- De Silva AD, Boesteanu A, Song R, Nagy N, Harhaj E, Harding CV, Joyce S. Thermolabile H-2Kb molecules expressed by transporter associated with antigen processing-deficient RMA-S cells are occupied by low-affinity peptides. J Immunol. 1999;163:4413–20. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
