

Review
doi: 10.1186/jbiol206.
Epub 2009 Dec 23.
The transcriptome of human monocyte subsets begins to emerge
Affiliations
- PMID: 20067595
- PMCID: PMC2804287
- DOI: 10.1186/jbiol206
Item in Clipboard
Review
The transcriptome of human monocyte subsets begins to emerge
J Biol.
2009.
Abstract
Human monocytes can be divided into subsets according to their expression or lack of the cell-surface antigen CD16. In papers published recently in the Journal of Proteome Research and in BMC Genomics, two groups publish independent transcriptome analyses of CD16(+) and CD16(-) monocytes, with revealing results.
Figures
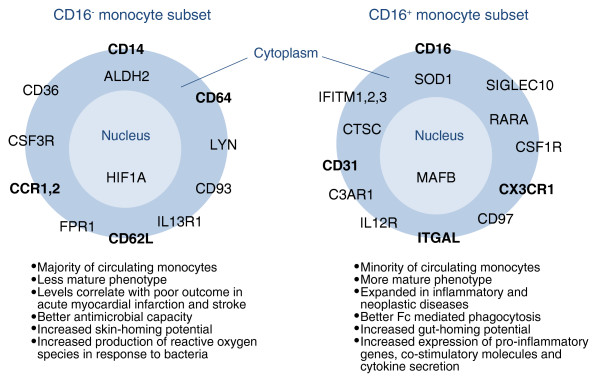
Summary of selected established and proposed functional differences between human CD16- and CD16+ monocyte subsets. Genes expressed at a high level in one subset but not in the other are indicated. Gene labels are positioned according to the location of the protein in the cell - in the plasma membrane, the cytoplasm or the nucleus. Several studies have previously confirmed high levels of expression in human CD16+ monocytes of the genes for the chemokine receptor CX3CR1, the integrin alpha chain ITGAL and the adhesion molecule CD31 and strong expression in human CD16- monocytes of genes for the adhesion molecule CD62L, the high-affinity Fc receptor for IgG (CD64), and the chemokine receptors CCR1 and CCR2. The new studies add many candidates to the monocyte subsets markers list, a selection of which is represented in the figure. Previously confirmed markers are in bold. A list of differentially expressed genes found by both [6,7] is given in Additional data file 1. Other abbreviations of markers: ALDH2, aldehyde dehydrogenase 2 family; C3AR1, complement component 3a receptor 1; CD26, dipeptidyl-peptidase 4; CD93, CD93 molecule; CD99, CD99 molecule; CSF1R, colony stimulating factor 1 receptor; CSF3R, colony stimulating factor 3 receptor; CTSC, cathepsin C; CD97, CD97 molecule; FPR1, formyl peptide receptor 1; HIF1A, hypoxia inducible factor 1, alpha subunit (basic helix-loop-helix transcription factor); IFITM1-3, Interferon-induced transmembrane protein 1, 2 and 3; IL12R, Interleukin 12 receptor; IL13R1, Interleukin 13 receptor 1, LYN, v-yes-1 Yamaguchi sarcoma viral related oncogene homolog; MAFB, v-maf musculoaponeurotic fibrosarcoma oncogene homolog B (avian); RARA, retinoic acid receptor alpha subunit; SIGLEC10, sialic acid binding Ig-like lectin 10; SOD1, soluble superoxide dismutase 1.
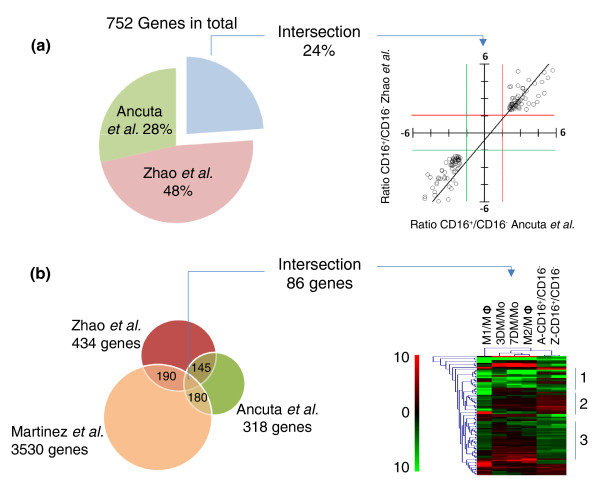
Direct comparison between high-throughput genomic studies hints at a complex interplay between genes as the basis for the differences between monocyte subsets. (a) The overlap between differentially expressed genes identified by Zhao et al. [6] and Ancuta et al. [7] was determined after eliminating all genes without gene symbols or duplicated. Merging the two studies we find that the number of genes differentially expressed between monocyte subsets amounts to 752 (100%). Of these 24% (145 genes) are genes detected in both studies; 48% of the remaining genes are detected as differentially expressed by [6] and 28% by [7]. A scatter-plot of the fold expression difference in CD16+ compared with CD16- monocytes shows correlation of the values in the two studies. Red and green cutoff lines extend along the values 1.5 and -1.5. The upper right quadrant and the lower left quadrant show genes with similar fold differences between the studies. (b) Overlap between the monocyte subset gene lists and a dataset of genes involved in human monocyte maturation induced by MCSF and macrophage activation induced by a combination of LPS and IFN-γ (M1) or IL-4 (M2) [8]. This comparison shows that 190 out of the 434 genes selected from [6], and 180 out of the 318 genes selected from [7], are also contained in the maturation/activation gene set. The overlap of the three lists amounts to 86 genes. A hierarchical clustering of these 86 genes shows that a proportion of them are regulated by MCSF and contrastingly regulated by the combination of IFN-γ and LPS used in [8] to drive macrophages towards classical activation. The differences in expression between subsets only partially correlate with the MCSF gene expression pattern. The tree can be divided in three main clusters (right). Cluster 1 shows genes downregulated by MCSF stimulation in total monocytes and whose levels are lower in CD16+ monocytes. Cluster 2 shows genes induced by MCSF stimulation that are highly expressed by CD16+ monocytes. These two clusters support the hypothesis that part of the differences between subsets correlates with an MCSF responsive phenotype for CD16+ monocytes. However, the behaviors of the genes in cluster 3 do not correlate with those of MCSF stimulation, and instead correlates with the inflammatory profile induced by the combination of IFN-γ and LPS used in [8] to drive macrophages towards classical activation. A list of the 86 overlapping genes is given in Additional data file 2. Other abbreviations used: M3, macrophage; Mo, monocytes; 3 DM, Monocytes stimulated for three days with MCSF equivalent to 3rd day macrophages; 7 DM, Monocytes stimulated for seven days with MCSF equivalent to 7th day macrophages. In the figure the ratio of RNA expression between given categories is indicated by the forward slash or stroke symbol "/". A-CD16+/CD16-, Ratio of RNA expression levels of CD16+ vs CD16- monocytes provided by Ancuta et al.[7]; Z-CD16+/CD16-, Ratio of CD16+ vs CD16- monocytes provided by Zhao et al.[6].
Similar articles
-
Human Monocyte Subset Distinctions and Function: Insights From Gene Expression Analysis.Front Immunol. 2020 Jun 4;11:1070. doi: 10.3389/fimmu.2020.01070. eCollection 2020. Front Immunol. 2020. PMID: 32582174 Free PMC article. Review.
-
Transcriptome analysis in patients with chronic kidney disease on hemodialysis disclosing a key role for CD16+CX3CR1+ monocytes.PLoS One. 2015 Apr 1;10(4):e0121750. doi: 10.1371/journal.pone.0121750. eCollection 2015. PLoS One. 2015. PMID: 25830914 Free PMC article.
-
Lower HDL-C levels are associated with higher expressions of CD16 on monocyte subsets in coronary atherosclerosis.Int J Med Sci. 2020 Aug 1;17(14):2171-2179. doi: 10.7150/ijms.47998. eCollection 2020. Int J Med Sci. 2020. PMID: 32922178 Free PMC article.
-
Standardized flow cytometry assay for identification of human monocytic heterogeneity and LRP1 expression in monocyte subpopulations: decreased expression of this receptor in nonclassical monocytes.Cytometry A. 2014 Jul;85(7):601-10. doi: 10.1002/cyto.a.22455. Epub 2014 Mar 17. Cytometry A. 2014. PMID: 24639232
-
The Role of Different Monocyte Subsets in the Pathogenesis of Atherosclerosis and Acute Coronary Syndromes.Scand J Immunol. 2015 Sep;82(3):163-73. doi: 10.1111/sji.12314. Scand J Immunol. 2015. PMID: 25997925 Review.
Cited by
-
Human Monocyte Subset Distinctions and Function: Insights From Gene Expression Analysis.Front Immunol. 2020 Jun 4;11:1070. doi: 10.3389/fimmu.2020.01070. eCollection 2020. Front Immunol. 2020. PMID: 32582174 Free PMC article. Review.
-
Pre-vaccination frequency of circulatory Tfh is associated with robust immune response to TV003 dengue vaccine.PLoS Pathog. 2022 Jan 21;18(1):e1009903. doi: 10.1371/journal.ppat.1009903. eCollection 2022 Jan. PLoS Pathog. 2022. PMID: 35061851 Free PMC article. Clinical Trial.
-
Macrophage heterogeneity in tissues: phenotypic diversity and functions.Immunol Rev. 2014 Nov;262(1):36-55. doi: 10.1111/imr.12223. Immunol Rev. 2014. PMID: 25319326 Free PMC article. Review.
-
Chronic Psoriatic Skin Inflammation Leads to Increased Monocyte Adhesion and Aggregation.J Immunol. 2015 Sep 1;195(5):2006-18. doi: 10.4049/jimmunol.1402307. Epub 2015 Jul 29. J Immunol. 2015. PMID: 26223654 Free PMC article.
-
Quaking promotes monocyte differentiation into pro-atherogenic macrophages by controlling pre-mRNA splicing and gene expression.Nat Commun. 2016 Mar 31;7:10846. doi: 10.1038/ncomms10846. Nat Commun. 2016. PMID: 27029405 Free PMC article.
References
-
- Passlick B, Flieger D, Ziegler-Heitbrock HW. Identification and characterization of a novel monocyte subpopulation in human peripheral blood. Blood. 1989;74:2527–2534. - PubMed
-
- Hanai H, Iida T, Takeuchi K, Watanabe F, Yamada M, Kikuyama M, Maruyama Y, Iwaoka Y, Hirayama K, Nagata S, Takai K. Adsorptive depletion of elevated proinflammatory CD14+CD16+DR++ monocytes in patients with inflammatory bowel disease. Am J Gastroenterol. 2008;103:1210–1216. doi: 10.1111/j.1572-0241.2007.01714.x. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
