

A novel small-molecule disrupts Stat3 SH2 domain-phosphotyrosine interactions and Stat3-dependent tumor processes
- PMID: 20067773
- PMCID: PMC3188443
- DOI: 10.1016/j.bcp.2010.01.001
A novel small-molecule disrupts Stat3 SH2 domain-phosphotyrosine interactions and Stat3-dependent tumor processes
Abstract
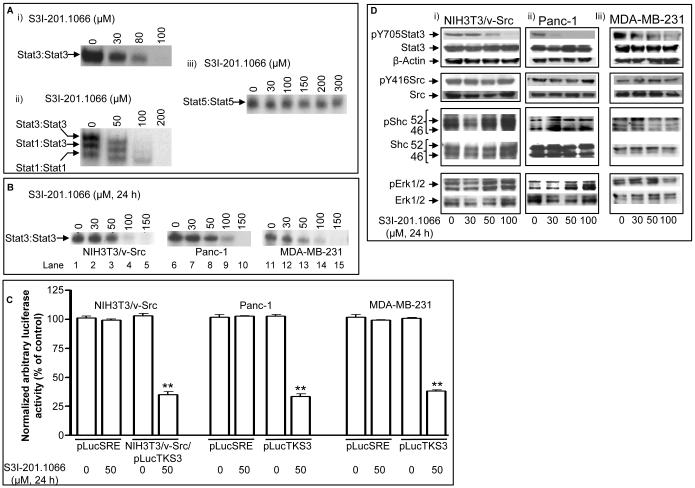
The molecular modeling of the phosphotyrosine (pTyr)-SH2 domain interaction in the Stat3:Stat3 dimerization, combined with in silico structural analysis of the Stat3 dimerization disruptor, S3I-201, has furnished a diverse set of analogs. We present evidence from in vitro biochemical and biophysical studies that the structural analog, S3I-201.1066 directly interacts with Stat3 or the SH2 domain, with an affinity (K(D)) of 2.74microM, and disrupts the binding of Stat3 to the cognate pTyr-peptide, GpYLPQTV-NH(2), with an IC(50) of 23microM. Moreover, S3I-201.1066 selectively blocks the association of Stat3 with the epidermal growth factor receptor (EGFR), and inhibits Stat3 tyrosine phosphorylation and nuclear translocation in EGF-stimulated mouse fibroblasts. In cancer cells that harbor aberrant Stat3 activity, S3I-201.1066 inhibits constitutive Stat3 DNA-binding and transcriptional activities. By contrast, S3I-201.1066 has no effect on Src activation or the EGFR-mediated activation of the Erk1/2(MAPK) pathway. S3I-201.1066 selectively suppresses the viability, survival, and malignant transformation of the human breast and pancreatic cancer lines and the v-Src-transformed mouse fibroblasts harboring persistently active Stat3. Treatment with S3I-201.1066 of malignant cells harboring aberrantly active Stat3 down-regulated the expression of c-Myc, Bcl-xL, Survivin, the matrix metalloproteinase 9, and VEGF. The in vivo administration of S3I-201.1066-induced significant antitumor response in mouse models of human breast cancer, which correlates with the inhibition of constitutively active Stat3 and the suppression of known Stat3-regulated genes. Our studies identify a novel small-molecule that binds with a high affinity to Stat3, blocks Stat3 activation and function, and thereby induces antitumor response in human breast tumor xenografts harboring persistently active Stat3.
2010 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
A novel inhibitor of STAT3 homodimerization selectively suppresses STAT3 activity and malignant transformation.Cancer Res. 2013 Mar 15;73(6):1922-33. doi: 10.1158/0008-5472.CAN-12-3175. Epub 2013 Jan 15. Cancer Res. 2013. PMID: 23322008 Free PMC article.
-
A cell-permeable Stat3 SH2 domain mimetic inhibits Stat3 activation and induces antitumor cell effects in vitro.J Biol Chem. 2010 Nov 12;285(46):35855-65. doi: 10.1074/jbc.M110.154088. Epub 2010 Aug 31. J Biol Chem. 2010. PMID: 20807764 Free PMC article.
-
Selective chemical probe inhibitor of Stat3, identified through structure-based virtual screening, induces antitumor activity.Proc Natl Acad Sci U S A. 2007 May 1;104(18):7391-6. doi: 10.1073/pnas.0609757104. Epub 2007 Apr 26. Proc Natl Acad Sci U S A. 2007. PMID: 17463090 Free PMC article.
-
STAT proteins as novel targets for cancer drug discovery.Expert Opin Ther Targets. 2004 Oct;8(5):409-22. doi: 10.1517/14728222.8.5.409. Expert Opin Ther Targets. 2004. PMID: 15469392 Review.
-
STAT3 as a target for inducing apoptosis in solid and hematological tumors.Cell Res. 2008 Feb;18(2):254-67. doi: 10.1038/cr.2008.18. Cell Res. 2008. PMID: 18227858 Free PMC article. Review.
Cited by
-
Orally bioavailable small-molecule inhibitor of transcription factor Stat3 regresses human breast and lung cancer xenografts.Proc Natl Acad Sci U S A. 2012 Jun 12;109(24):9623-8. doi: 10.1073/pnas.1121606109. Epub 2012 May 23. Proc Natl Acad Sci U S A. 2012. PMID: 22623533 Free PMC article.
-
STAT3 activity is necessary and sufficient for the development of immune-mediated myocarditis in mice and promotes progression to dilated cardiomyopathy.EMBO Mol Med. 2013 Apr;5(4):572-90. doi: 10.1002/emmm.201201876. Epub 2013 Mar 5. EMBO Mol Med. 2013. PMID: 23460527 Free PMC article.
-
Docosahexaenoic Acid Inhibits Cell Proliferation through a Suppression of c-Myc Protein in Pancreatic Ductal Adenocarcinoma Cells.Antioxidants (Basel). 2021 Oct 28;10(11):1721. doi: 10.3390/antiox10111721. Antioxidants (Basel). 2021. PMID: 34829591 Free PMC article.
-
Linker Variation and Structure-Activity Relationship Analyses of Carboxylic Acid-based Small Molecule STAT3 Inhibitors.ACS Med Chem Lett. 2018 Feb 16;9(3):250-255. doi: 10.1021/acsmedchemlett.7b00544. eCollection 2018 Mar 8. ACS Med Chem Lett. 2018. PMID: 29541369 Free PMC article.
-
Hepcidin antagonists for potential treatments of disorders with hepcidin excess.Front Pharmacol. 2014 Apr 28;5:86. doi: 10.3389/fphar.2014.00086. eCollection 2014. Front Pharmacol. 2014. PMID: 24808863 Free PMC article. Review.
References
-
- Darnell JE., Jr. Transcription factors as targets for cancer therapy. Nat Rev Cancer. 2002;2:740–9. - PubMed
-
- Schröder M, Kroeger K, Volk HD, Eidne KA, Grütz G. Preassociation of nonactivated STAT3 molecules demonstrated in living cells using bioluminescence resonance energy transfer: a new model of STAT activation. J Leukoc Biol. 2004;75:792–7. - PubMed
-
- Bromberg J, Darnell JE., Jr. The role of STATs in transcriptional control and their impact on cellular function. Oncogene. 2000;19:2468–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
