

Cytoskeletal coherence requires myosin-IIA contractility
- PMID: 20067993
- PMCID: PMC2816186
- DOI: 10.1242/jcs.058297
Cytoskeletal coherence requires myosin-IIA contractility
Abstract
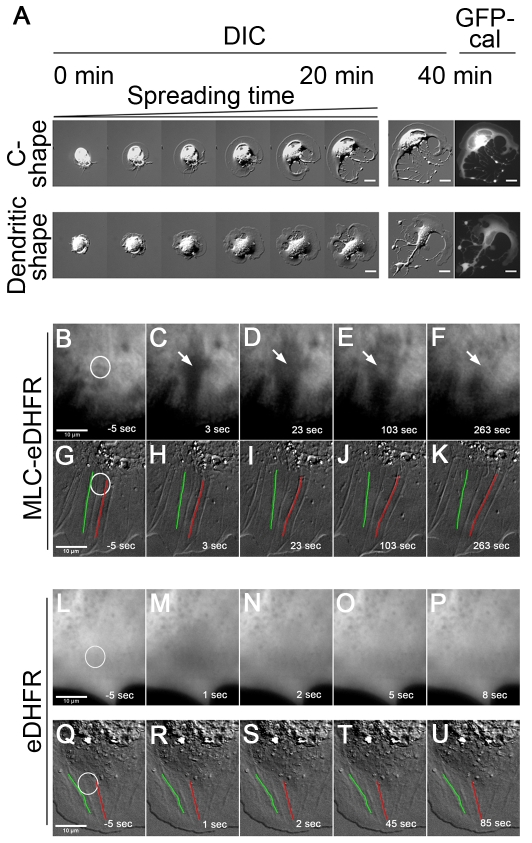
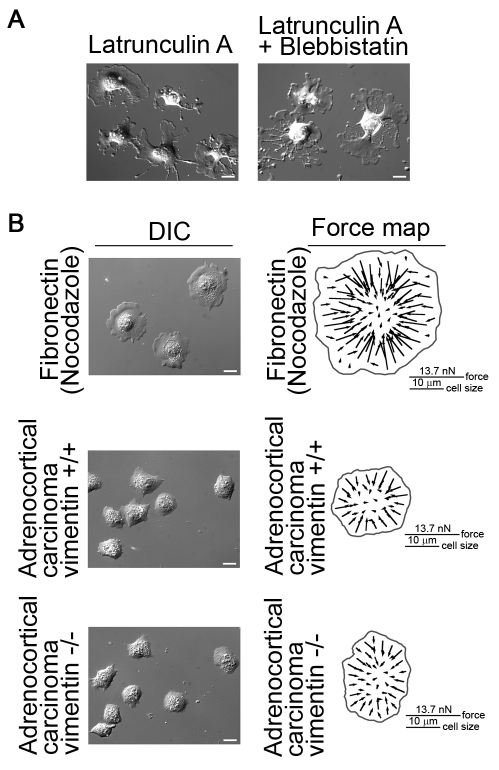
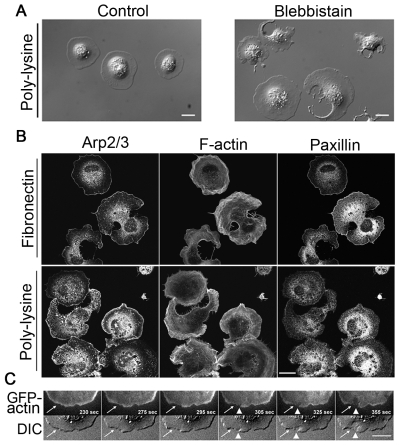
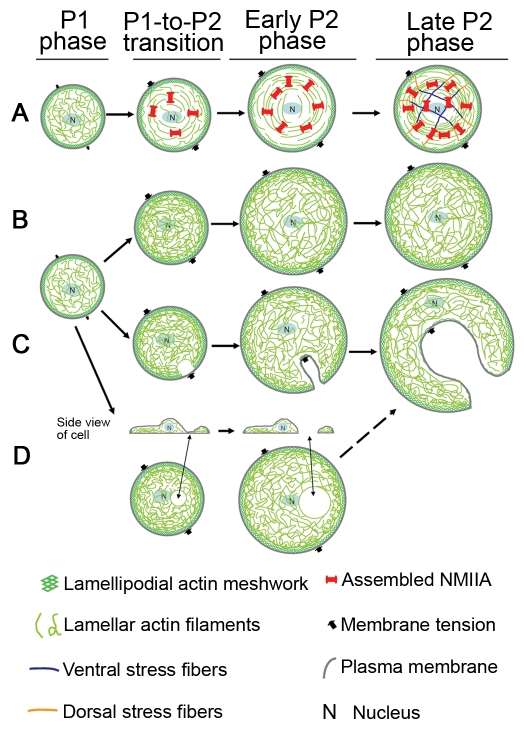
Maintaining a physical connection across cytoplasm is crucial for many biological processes such as matrix force generation, cell motility, cell shape and tissue development. However, in the absence of stress fibers, the coherent structure that transmits force across the cytoplasm is not understood. We find that nonmuscle myosin-II (NMII) contraction of cytoplasmic actin filaments establishes a coherent cytoskeletal network irrespective of the nature of adhesive contacts. When NMII activity is inhibited during cell spreading by Rho kinase inhibition, blebbistatin, caldesmon overexpression or NMIIA RNAi, the symmetric traction forces are lost and cell spreading persists, causing cytoplasm fragmentation by membrane tension that results in 'C' or dendritic shapes. Moreover, local inactivation of NMII by chromophore-assisted laser inactivation causes local loss of coherence. Actin filament polymerization is also required for cytoplasmic coherence, but microtubules and intermediate filaments are dispensable. Loss of cytoplasmic coherence is accompanied by loss of circumferential actin bundles. We suggest that NMIIA creates a coherent actin network through the formation of circumferential actin bundles that mechanically link elements of the peripheral actin cytoskeleton where much of the force is generated during spreading.
Figures



References
-
- Alexandrova A. Y., Dugina V. B., Ivanova O. Y., Kaverina I. N., Vasiliev J. M. (1998). Scatter factor induces segregation of multinuclear cells into several discrete motile domains. Cell Motil. Cytoskeleton 39, 147-158 - PubMed
-
- Balaban N. Q., Schwarz U. S., Riveline D., Goichberg P., Tzur G., Sabanay I., Mahalu D., Safran S., Bershadsky A., Addadi L., et al. (2001). Force and focal adhesion assembly: a close relationship studied using elastic micropatterned substrates. Nat. Cell Biol. 3, 466-472 - PubMed
-
- Bao J., Ma X., Liu C., Adelstein R. S. (2007). Replacement of nonmuscle myosin II-B with II-A rescues brain but not cardiac defects in mice. J. Biol. Chem. 282, 22102-22111 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
