

Taking the plunge: integrating structural, enzymatic and computational insights into a unified model for membrane-immersed rhomboid proteolysis
- PMID: 20070259
- PMCID: PMC3131108
- DOI: 10.1042/BJ20090861
Taking the plunge: integrating structural, enzymatic and computational insights into a unified model for membrane-immersed rhomboid proteolysis
Abstract
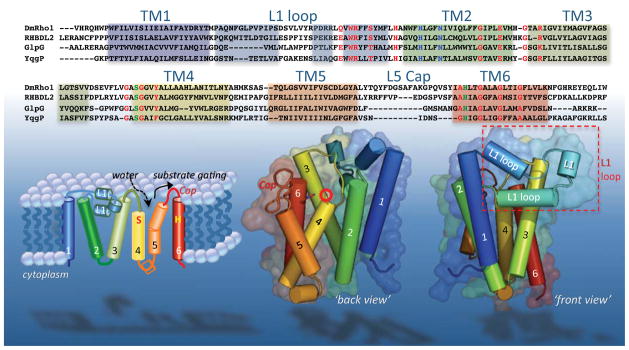
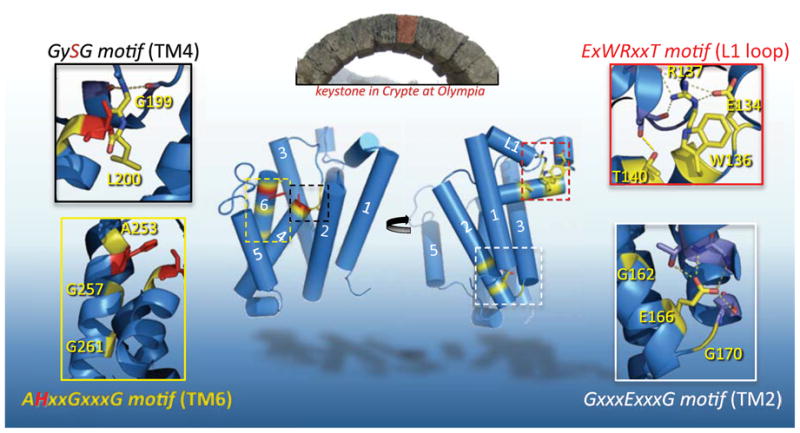
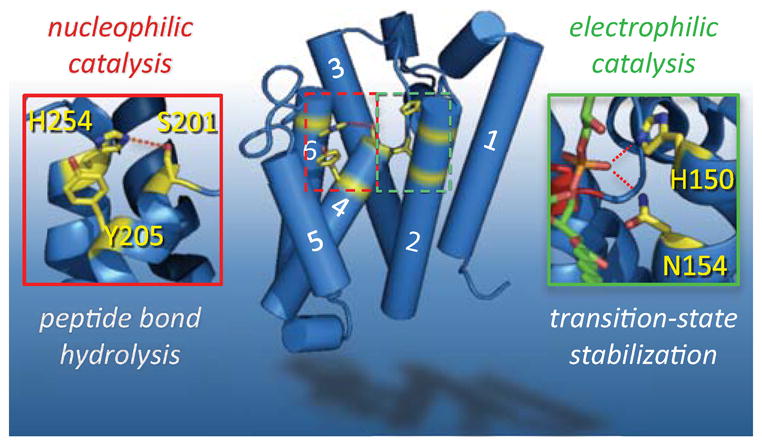
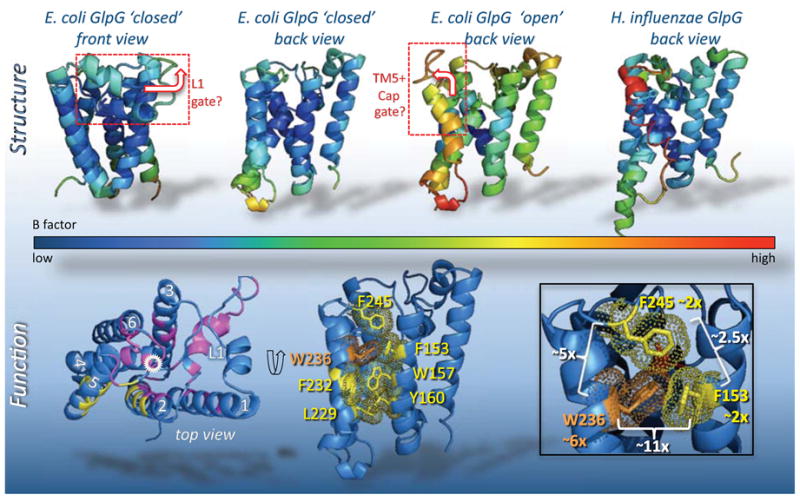
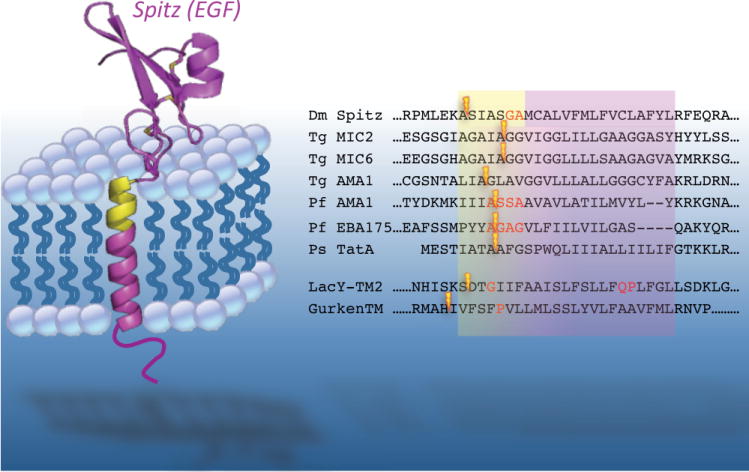
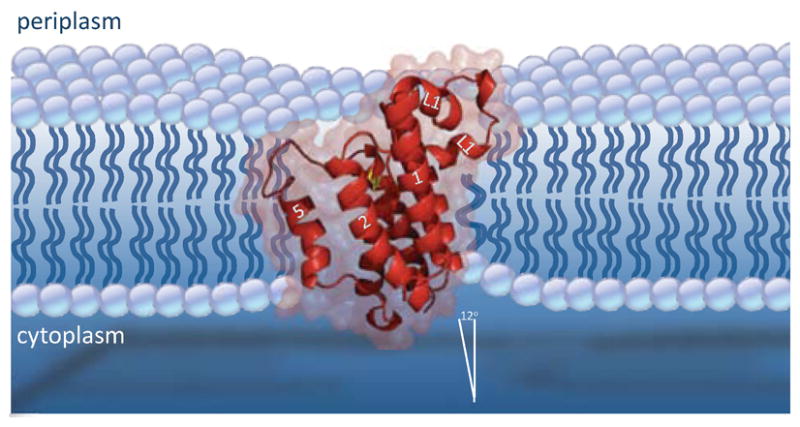
Rhomboid proteases are a fascinating class of enzymes that combine a serine protease active site within the core of an integral membrane protein. Despite having key roles in animal cell signalling and microbial pathogenesis, the membrane-immersed nature of these enzymes had long imposed obstacles to elucidating their biochemical mechanisms. But recent multidisciplinary approaches, including eight crystal structures, four computer simulations and nearly 100 engineered mutants interrogated in vivo and in vitro, are coalescing into an integrated model for one rhomboid orthologue in particular, bacterial GlpG. The protein creates a central hydrated microenvironment immersed below the membrane surface to support hydrolysis by its serine protease-like catalytic apparatus. Four conserved architectural elements in particular act as 'keystones' to stabilize this structure, and the lateral membrane-embedded L1 loop functions as a 'flotation device' to position the protease tilted in the membrane. Complex interplay between lateral substrate gating by rhomboid, substrate unwinding and local membrane thinning leads to intramembrane proteolysis of selected target proteins. Although far from complete, studies with GlpG currently offer the best prospect for achieving a thorough and sophisticated understanding of a simplified intramembrane protease.
Figures
References
-
- Lopez-Otin C, Overall CM. Protease degradomics: a new challenge for proteomics. Nat Rev Mol Cell Biol. 2002;3:509–519. - PubMed
-
- Blow DM, Birktoft JJ, Hartley BS. Role of a buried acid group in the mechanism of action of chymotrypsin. Nature. 1969;221:337–340. - PubMed
-
- Blow D. So do we understand how enzymes work? Structure. 2000;8:R77–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases