

Myosin 1G is an abundant class I myosin in lymphocytes whose localization at the plasma membrane depends on its ancient divergent pleckstrin homology (PH) domain (Myo1PH)
- PMID: 20071333
- PMCID: PMC2838290
- DOI: 10.1074/jbc.M109.086959
Myosin 1G is an abundant class I myosin in lymphocytes whose localization at the plasma membrane depends on its ancient divergent pleckstrin homology (PH) domain (Myo1PH)
Abstract
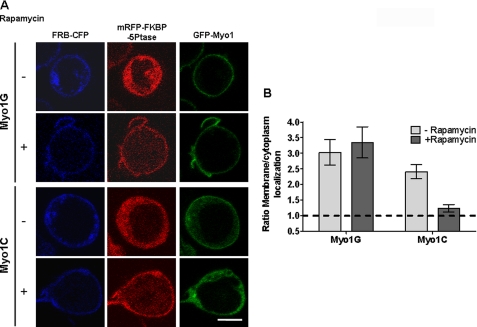
Class I myosins, which link F-actin to membrane, are largely undefined in lymphocytes. Mass spectrometric analysis of lymphocytes identified two short tail forms: (Myo1G and Myo1C) and one long tail (Myo1F). We investigated Myo1G, the most abundant in T-lymphocytes, and compared key findings with Myo1C and Myo1F. Myo1G localizes to the plasma membrane and associates in an ATP-releasable manner to the actin-containing insoluble pellet. The IQ+tail region of Myo1G (Myo1C and Myo1F) is sufficient for membrane localization, but membrane localization is augmented by the motor domain. The minimal region lacks IQ motifs but includes: 1) a PH-like domain; 2) a "Pre-PH" region; and 3) a "Post-PH" region. The Pre-PH predicted alpha helices may contribute electrostatically, because two conserved basic residues on one face are required for optimal membrane localization. Our sequence analysis characterizes the divergent PH domain family, Myo1PH, present also in long tail myosins, in eukaryotic proteins unrelated to myosins, and in a probable ancestral protein in prokaryotes. The Myo1G Myo1PH domain utilizes the classic lipid binding site for membrane association, because mutating either of two basic residues in the "signature motif" destroys membrane localization. Mutation of each basic residue of the Myo1G Myo1PH domain reveals another critical basic residue in the beta3 strand, which is shared only by Myo1D. Myo1G differs from Myo1C in its phosphatidylinositol 4,5-bisphosphate dependence for membrane association, because membrane localization of phosphoinositide 5-phosphatase releases Myo1C from the membrane but not Myo1G. Thus Myo1PH domains likely play universal roles in myosin I membrane association, but different isoforms have diverged in their binding specificity.
Figures








Similar articles
-
Myo1c binds phosphoinositides through a putative pleckstrin homology domain.Mol Biol Cell. 2006 Nov;17(11):4856-65. doi: 10.1091/mbc.e06-05-0449. Epub 2006 Sep 13. Mol Biol Cell. 2006. PMID: 16971510 Free PMC article.
-
Selective localization of myosin-I proteins in macropinosomes and actin waves.Cytoskeleton (Hoboken). 2016 Feb;73(2):68-82. doi: 10.1002/cm.21275. Epub 2016 Feb 22. Cytoskeleton (Hoboken). 2016. PMID: 26801966 Free PMC article.
-
Myosin-X, a novel myosin with pleckstrin homology domains, associates with regions of dynamic actin.J Cell Sci. 2000 Oct;113 Pt 19:3439-51. doi: 10.1242/jcs.113.19.3439. J Cell Sci. 2000. PMID: 10984435
-
Long-Tailed Unconventional Class I Myosins in Health and Disease.Int J Mol Sci. 2020 Apr 7;21(7):2555. doi: 10.3390/ijms21072555. Int J Mol Sci. 2020. PMID: 32272642 Free PMC article. Review.
-
Pleckstrin homology domains: not just for phosphoinositides.Biochem Soc Trans. 2004 Nov;32(Pt 5):707-11. doi: 10.1042/BST0320707. Biochem Soc Trans. 2004. PMID: 15493994 Review.
Cited by
-
Membrane-bound myo1c powers asymmetric motility of actin filaments.Curr Biol. 2012 Sep 25;22(18):1688-92. doi: 10.1016/j.cub.2012.06.069. Epub 2012 Aug 2. Curr Biol. 2012. PMID: 22863317 Free PMC article.
-
Myo1e binds anionic phospholipids with high affinity.Biochemistry. 2010 Nov 2;49(43):9353-60. doi: 10.1021/bi1012657. Biochemistry. 2010. PMID: 20860408 Free PMC article.
-
MYO1D binds with kinase domain of the EGFR family to anchor them to plasma membrane before their activation and contributes carcinogenesis.Oncogene. 2019 Dec;38(49):7416-7432. doi: 10.1038/s41388-019-0954-8. Epub 2019 Aug 16. Oncogene. 2019. PMID: 31420606
-
Identification of novel families and classification of the C2 domain superfamily elucidate the origin and evolution of membrane targeting activities in eukaryotes.Gene. 2010 Dec 1;469(1-2):18-30. doi: 10.1016/j.gene.2010.08.006. Epub 2010 Aug 14. Gene. 2010. PMID: 20713135 Free PMC article.
-
Molecular basis of dynamic relocalization of Dictyostelium myosin IB.J Biol Chem. 2012 Apr 27;287(18):14923-36. doi: 10.1074/jbc.M111.318667. Epub 2012 Feb 24. J Biol Chem. 2012. PMID: 22367211 Free PMC article.
References
-
- Jacobelli J., Chmura S. A., Buxton D. B., Davis M. M., Krummel M. F. (2004) Nat. Immunol. 5, 531–538 - PubMed
-
- Richards T. A., Cavalier-Smith T. (2005) Nature 436, 1113–1118 - PubMed
-
- Kalhammer G., Bähler M. (2000) Essays Biochem. 35, 33–42 - PubMed
-
- Engqvist-Goldstein A. E., Drubin D. G. (2003) Annu. Rev. Cell Dev. Biol. 19, 287–332 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
