

The Hsp90 cochaperone, FKBP51, increases Tau stability and polymerizes microtubules
- PMID: 20071522
- PMCID: PMC2830818
- DOI: 10.1523/JNEUROSCI.4815-09.2010
The Hsp90 cochaperone, FKBP51, increases Tau stability and polymerizes microtubules
Abstract
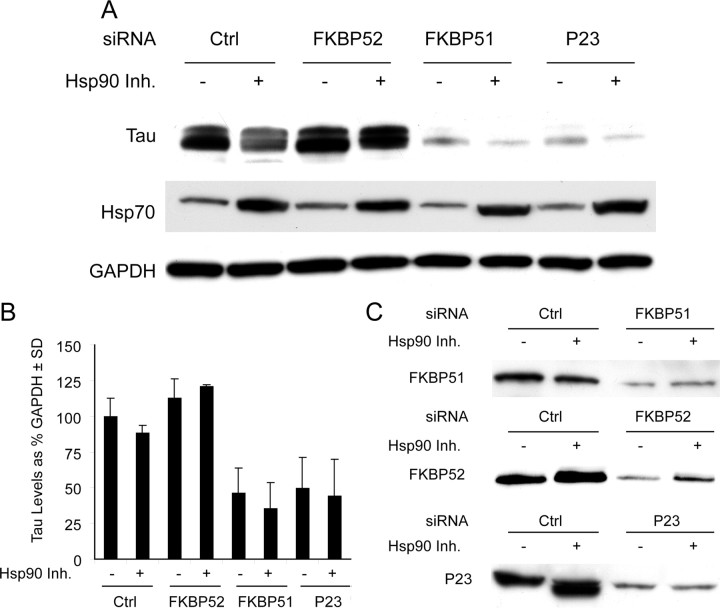
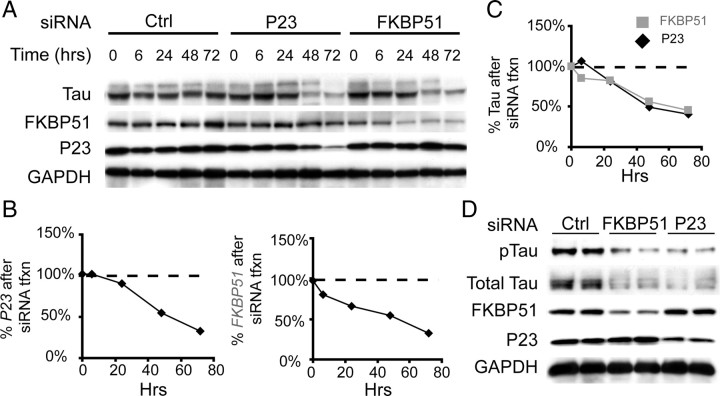
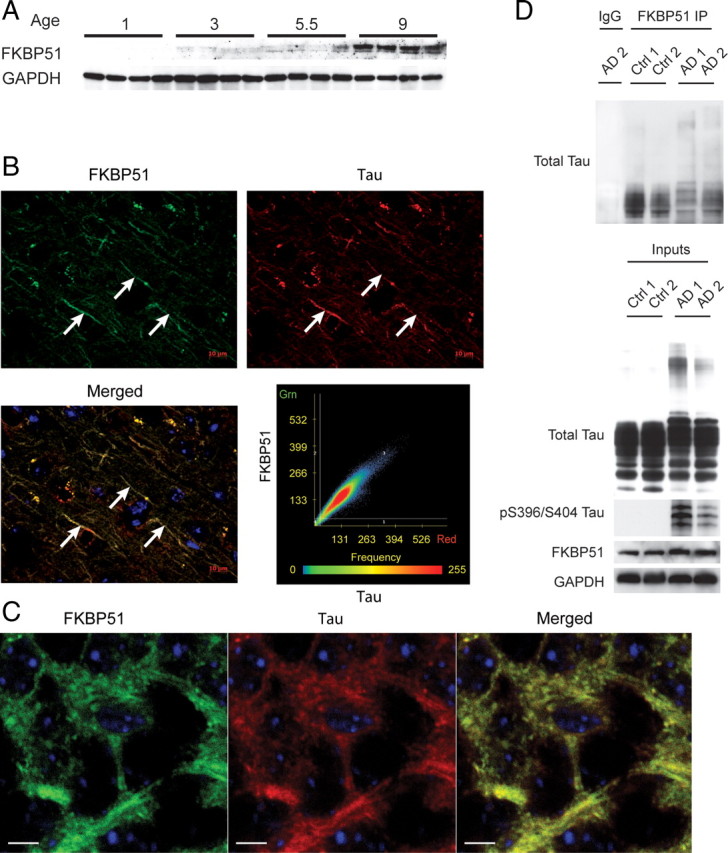
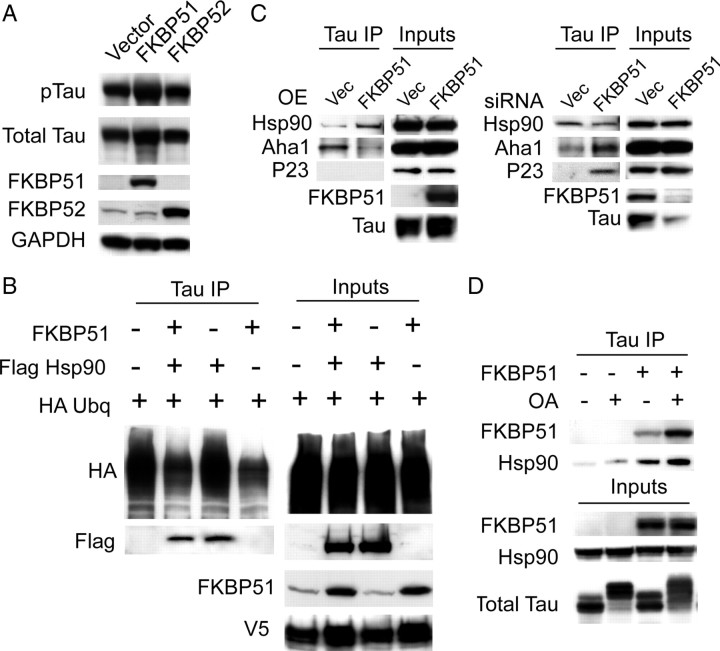
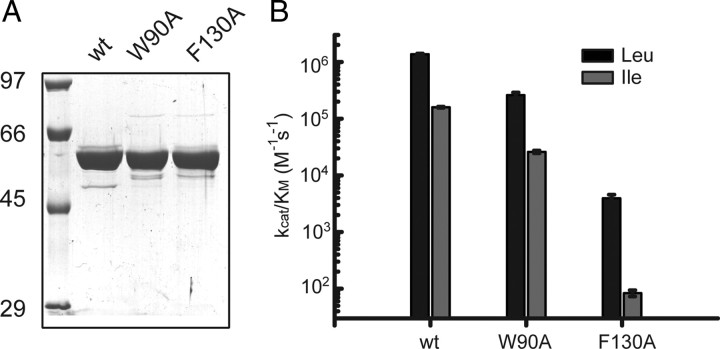
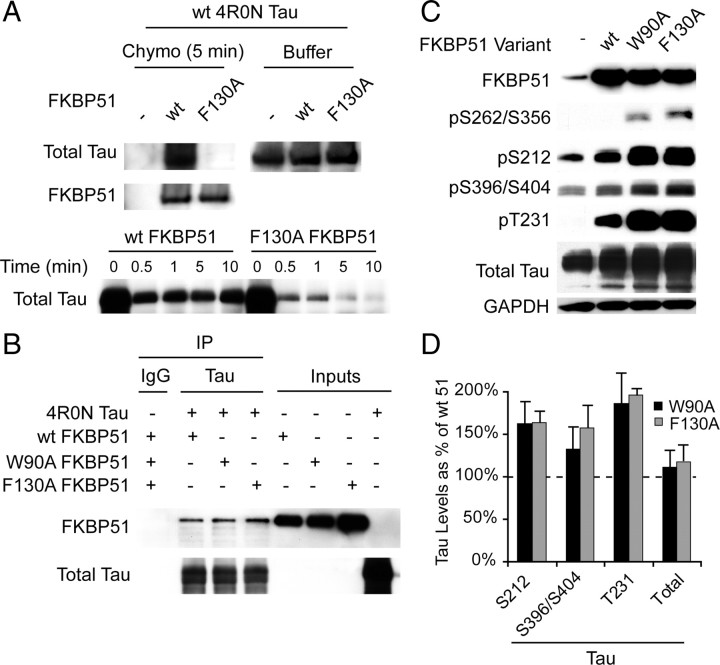
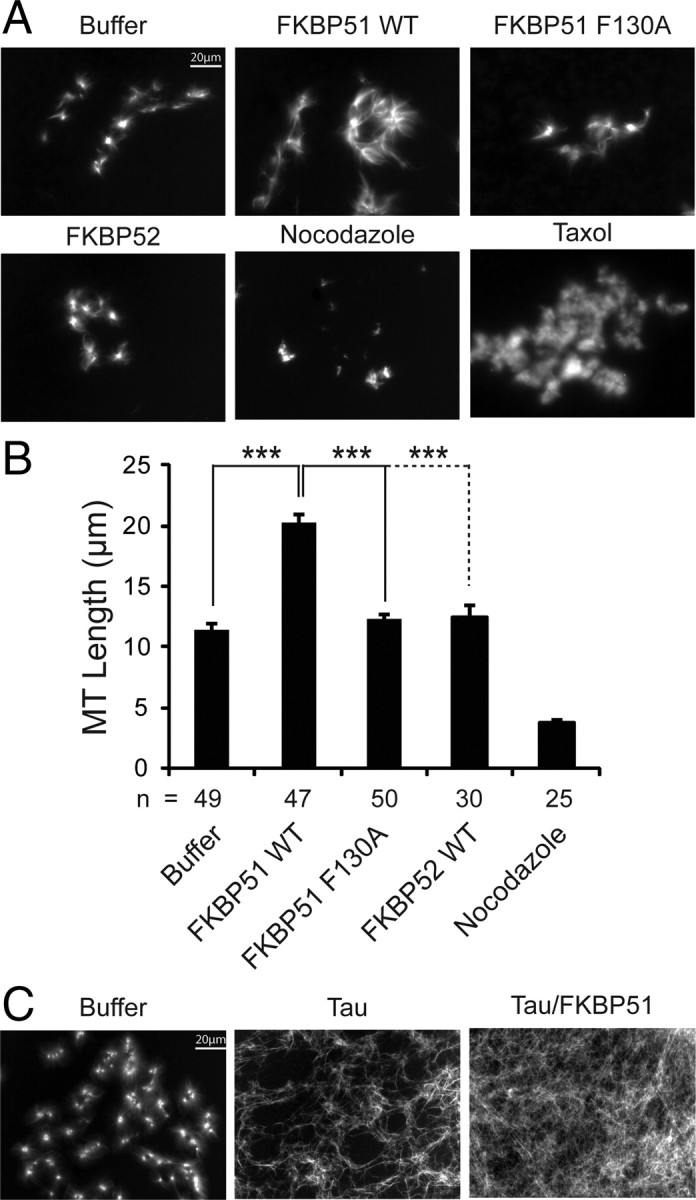
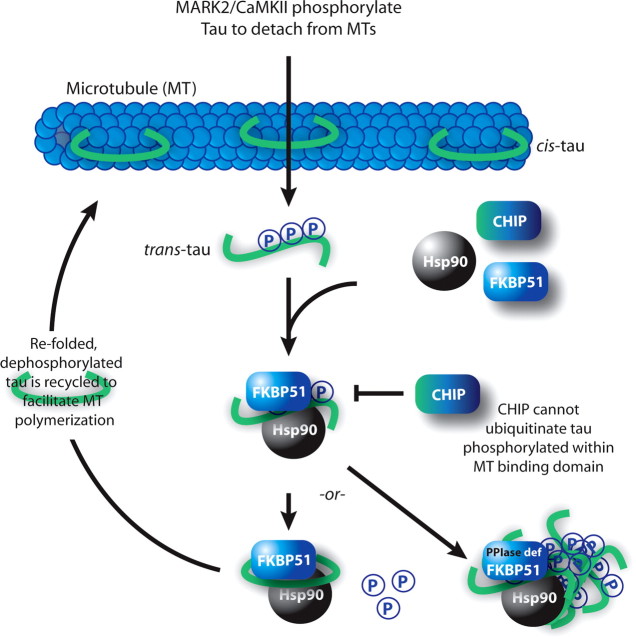
Imbalanced protein load within cells is a critical aspect for most diseases of aging. In particular, the accumulation of proteins into neurotoxic aggregates is a common thread for a host of neurodegenerative diseases. Our previous work demonstrated that age-related changes to the cellular chaperone repertoire contributes to abnormal buildup of the microtubule-associated protein tau that accumulates in a group of diseases termed tauopathies, the most common being Alzheimer's disease. Here, we show that the Hsp90 cochaperone, FK506-binding protein 51 (FKBP51), which possesses both an Hsp90-interacting tetratricopeptide domain and a peptidyl-prolyl cis-trans isomerase (PPIase) domain, prevents tau clearance and regulates its phosphorylation status. Regulation of the latter is dependent on the PPIase activity of FKBP51. FKB51 enhances the association of tau with Hsp90, but the FKBP51/tau interaction is not dependent on Hsp90. In vitro FKBP51 stabilizes microtubules with tau in a reaction depending on the PPIase activity of FKBP51. Based on these new findings, we propose that FKBP51 can use the Hsp90 complex to isomerize tau, altering its phosphorylation pattern and stabilizing microtubules.
Figures
Similar articles
-
Accelerated neurodegeneration through chaperone-mediated oligomerization of tau.J Clin Invest. 2013 Oct;123(10):4158-69. doi: 10.1172/JCI69003. Epub 2013 Sep 3. J Clin Invest. 2013. PMID: 23999428 Free PMC article.
-
Structure and pro-toxic mechanism of the human Hsp90/PPIase/Tau complex.Nat Commun. 2018 Oct 31;9(1):4532. doi: 10.1038/s41467-018-06880-0. Nat Commun. 2018. PMID: 30382094 Free PMC article.
-
The role of the FKBP51-Hsp90 complex in Alzheimer's disease: An emerging new drug target.Cell Stress Chaperones. 2024 Dec;29(6):792-804. doi: 10.1016/j.cstres.2024.11.006. Epub 2024 Nov 29. Cell Stress Chaperones. 2024. PMID: 39615785 Free PMC article. Review.
-
FKBP51 functions in the regulation of circadian rhythm and Alzheimer's disease.Cell Stress Chaperones. 2025 Mar;30(2):81-83. doi: 10.1016/j.cstres.2025.02.002. Epub 2025 Feb 9. Cell Stress Chaperones. 2025. PMID: 39933601 Free PMC article. Review.
-
The structure of an Hsp90-immunophilin complex reveals cochaperone recognition of the client maturation state.Mol Cell. 2021 Sep 2;81(17):3496-3508.e5. doi: 10.1016/j.molcel.2021.07.023. Epub 2021 Aug 10. Mol Cell. 2021. PMID: 34380015 Free PMC article.
Cited by
-
Hsp90-binding immunophilin FKBP51 forms complexes with hTERT enhancing telomerase activity.Mol Oncol. 2016 Aug;10(7):1086-98. doi: 10.1016/j.molonc.2016.05.002. Epub 2016 May 17. Mol Oncol. 2016. Retraction in: Mol Oncol. 2024 Aug;18(8):2060. doi: 10.1002/1878-0261.13676. PMID: 27233944 Free PMC article. Retracted.
-
Tau Protein Squired by Molecular Chaperones During Alzheimer's Disease.J Mol Neurosci. 2018 Nov;66(3):356-368. doi: 10.1007/s12031-018-1174-3. Epub 2018 Sep 28. J Mol Neurosci. 2018. PMID: 30267382 Review.
-
Chaperone-mediated autophagy in neurodegenerative diseases: mechanisms and therapy.Mol Cell Biochem. 2023 Oct;478(10):2173-2190. doi: 10.1007/s11010-022-04640-9. Epub 2023 Jan 25. Mol Cell Biochem. 2023. PMID: 36695937 Review.
-
Hsp90 and Its Co-Chaperones in Neurodegenerative Diseases.Int J Mol Sci. 2019 Oct 9;20(20):4976. doi: 10.3390/ijms20204976. Int J Mol Sci. 2019. PMID: 31600883 Free PMC article. Review.
-
Mapping interactions with the chaperone network reveals factors that protect against tau aggregation.Nat Struct Mol Biol. 2018 May;25(5):384-393. doi: 10.1038/s41594-018-0057-1. Epub 2018 Apr 30. Nat Struct Mol Biol. 2018. PMID: 29728653 Free PMC article.
References
-
- Auluck PK, Chan HY, Trojanowski JQ, Lee VM, Bonini NM. Chaperone suppression of alpha-synuclein toxicity in a Drosophila model for Parkinson's disease. Science. 2002;295:865–868. - PubMed
-
- Blatch GL, Lässle M. The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays. 1999;21:932–939. - PubMed
-
- Budde PP, Desai A, Heald R. Analysis of microtubule polymerization in vitro and during the cell cycle in Xenopus egg extracts. Methods. 2006;38:29–34. - PubMed
-
- Chambraud B, Belabes H, Fontaine-Lenoir V, Fellous A, Baulieu EE. The immunophilin FKBP52 specifically binds to tubulin and prevents microtubule formation. FASEB J. 2007;21:2787–2797. - PubMed
-
- Cox MB, Smith DF. Functions of the Hsp90-binding FKBP immunophilin. In: Blatch GL, editor. Networking of chaperones by co-chaperones. Austin, TX: Landes Bioscience/Eurekah.com; 2006. pp. 13–23.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous