

De novo neurogenesis in adult hypothalamus as a compensatory mechanism to regulate energy balance
- PMID: 20071537
- PMCID: PMC3080014
- DOI: 10.1523/JNEUROSCI.2479-09.2010
De novo neurogenesis in adult hypothalamus as a compensatory mechanism to regulate energy balance
Abstract
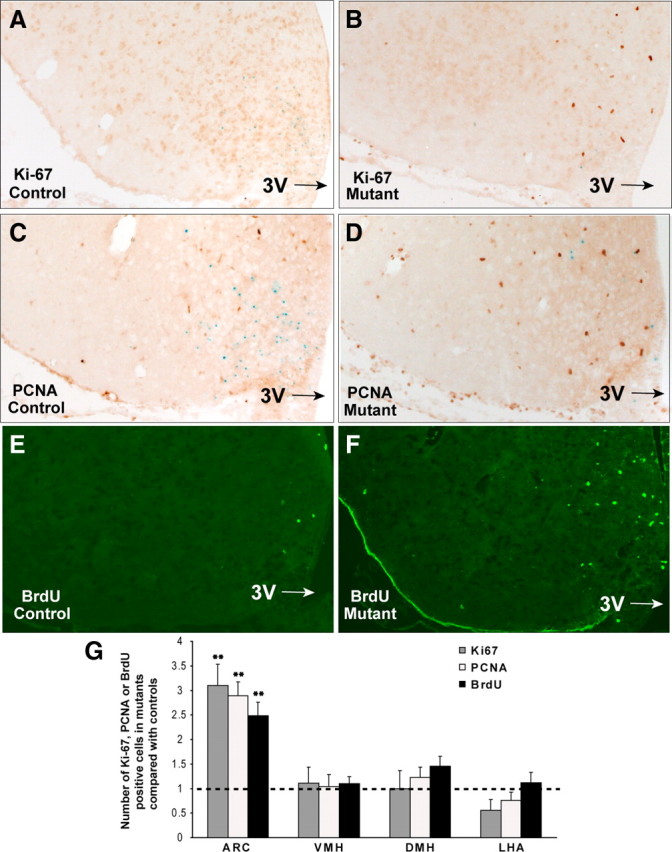
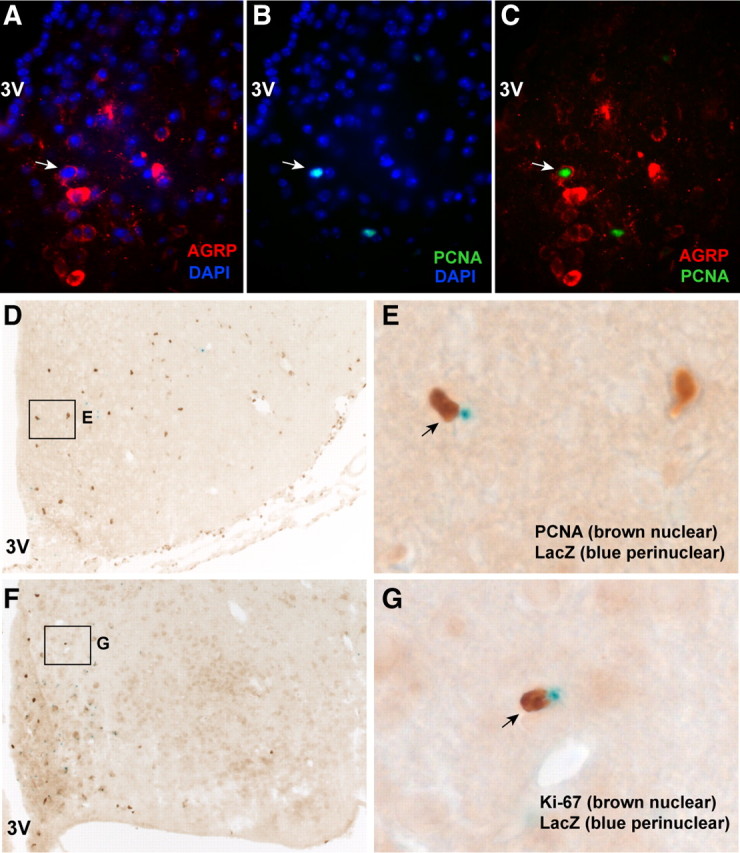
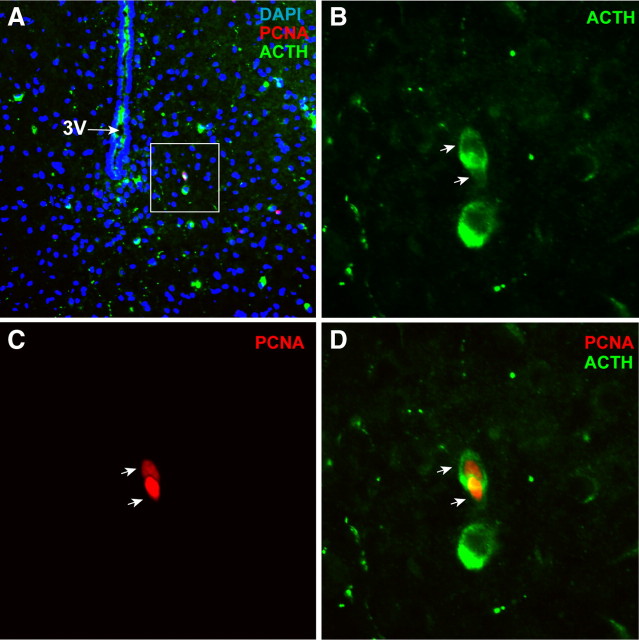
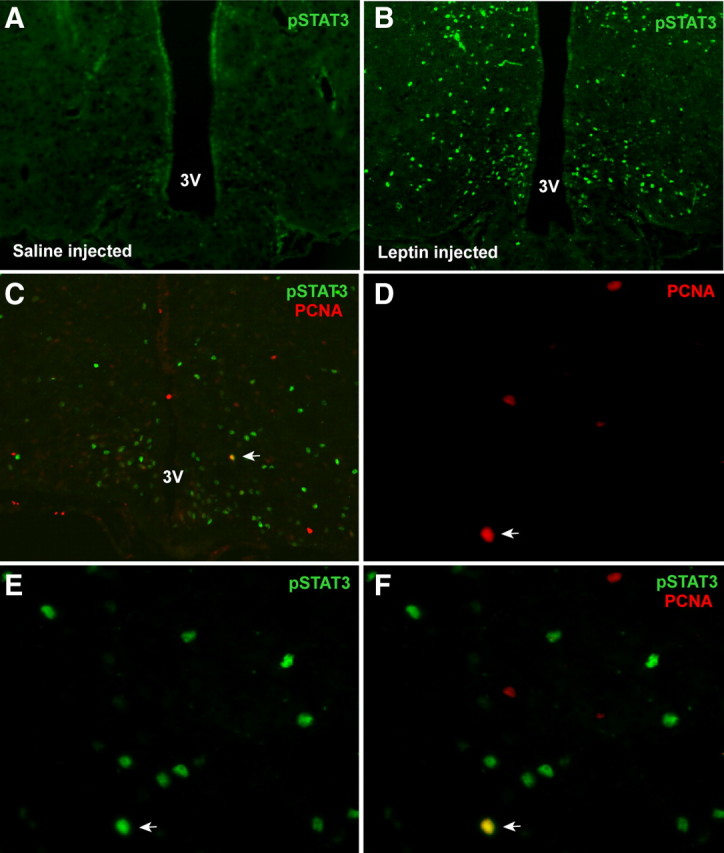
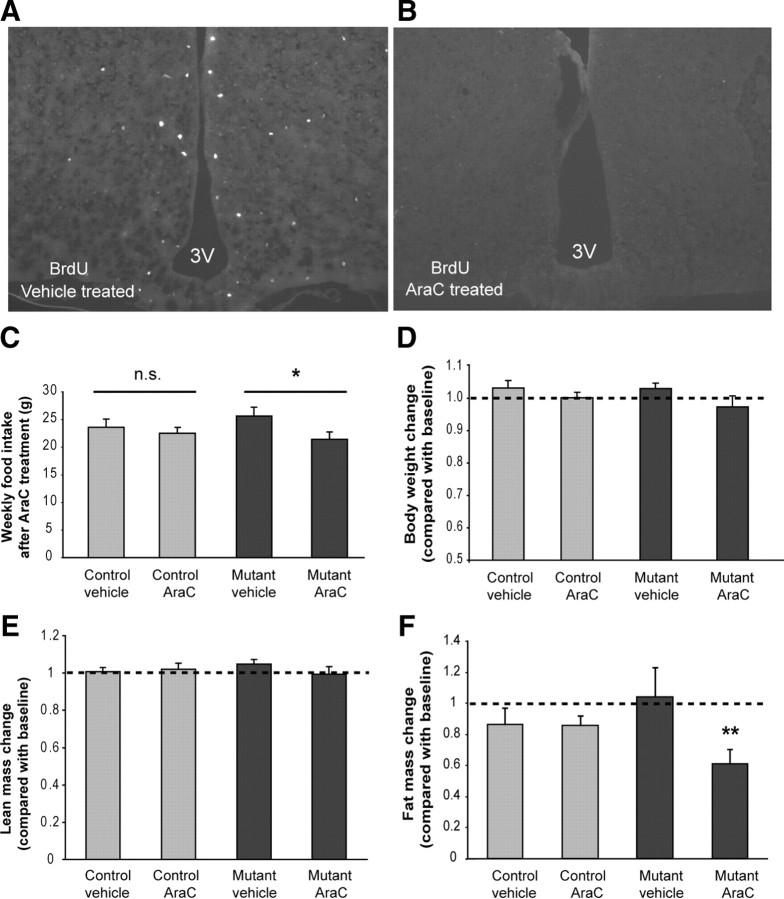
The ability to develop counter-regulatory mechanisms to maintain energy balance in response to environmental and physiologic insults is essential for survival, but the mechanisms underlying these compensatory regulations are poorly understood. Agouti-related peptide (AGRP) and Neuropeptide Y are potent orexigens and are coexpressed in neurons in the arcuate nucleus of the hypothalamus. Acute ablation of these neurons leads to severe anorexia and weight loss, whereas progressive degeneration of these neurons has minimal impact on food intake and body weight, suggesting that compensatory mechanisms are developed to maintain orexigenic drive. In this study, we show that cell proliferation is increased in the hypothalamus of adult mutant animals in which AgRP neurons undergo progressive neurodegeneration due to deletion of mitochondrial transcription factor A, and that a subset of these newly generated cells differentiate into AgRP neurons along with other resident neuronal subtypes. Furthermore, some of the newly generated cells are capable of responding to leptin, and a central blockade of cell proliferation in adult animals results in decreases in food intake and body adiposity in mutant but not in control animals. Our study indicates that neurons important for energy homeostasis can be regenerated in adult feeding centers under neurodegenerative conditions. It further suggests that de novo neurogenesis might serve as a compensatory mechanism contributing to the plastic control of energy balance in response to environmental and physiologic insults.
Figures
References
-
- Abdipranoto A, Wu S, Stayte S, Vissel B. The role of neurogenesis in neurodegenerative diseases and its implications for therapeutic development. CNS Neurol Disord Drug Targets. 2008;7:187–210. - PubMed
-
- Abercrombie M. Estimation of nuclear population from microtome sections. Anat Rec. 1946;94:239–247. - PubMed
-
- Cowley MA, Pronchuk N, Fan W, Dinulescu DM, Colmers WF, Cone RD. Integration of NPY, AGRP, and melanocortin signals in the hypothalamic paraventricular nucleus: evidence of a cellular basis for the adipostat. Neuron. 1999;24:155–163. - PubMed
-
- Cowley MA, Smart JL, Rubinstein M, Cerdan MG, Diano S, Horvath TL, Cone RD, Low MJ. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature. 2001;411:480–484. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases