

Measles virus selectively blind to signaling lymphocytic activation molecule (SLAM; CD150) is attenuated and induces strong adaptive immune responses in rhesus monkeys
- PMID: 20071568
- PMCID: PMC2838096
- DOI: 10.1128/JVI.02304-09
Measles virus selectively blind to signaling lymphocytic activation molecule (SLAM; CD150) is attenuated and induces strong adaptive immune responses in rhesus monkeys
Abstract
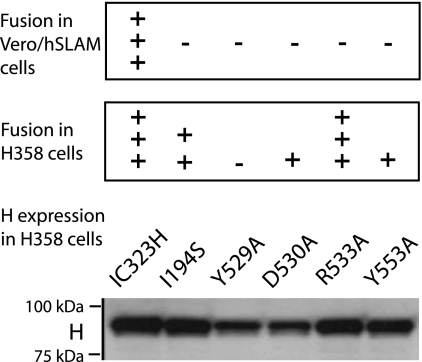
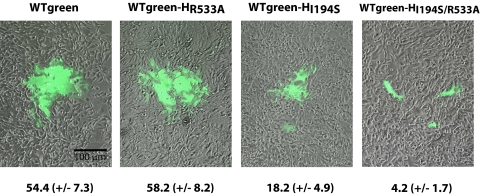
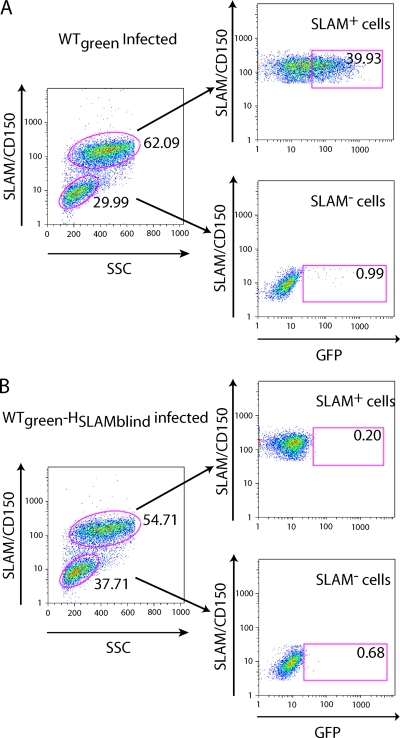
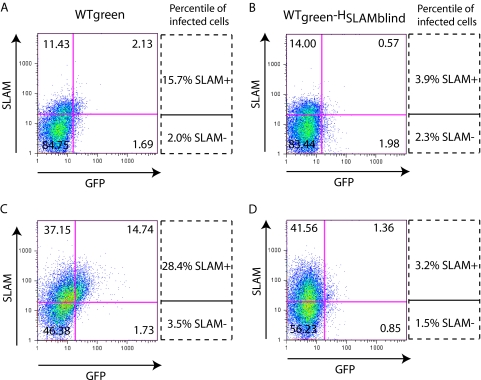
The signaling lymphocytic activation molecule (SLAM; CD150) is the immune cell receptor for measles virus (MV). To assess the importance of the SLAM-MV interactions for virus spread and pathogenesis, we generated a wild-type IC-B MV selectively unable to recognize human SLAM (SLAM-blind). This virus differs from the fully virulent wild-type IC-B strain by a single arginine-to-alanine substitution at amino acid 533 of the attachment protein hemagglutinin and infects cells through SLAM about 40 times less efficiently than the isogenic wild-type strain. Ex vivo, this virus infects primary lymphocytes at low levels regardless of SLAM expression. When a group of six rhesus monkeys (Macaca mulatta) was inoculated intranasally with the SLAM-blind virus, no clinical symptoms were documented. Only one monkey had low-level viremia early after infection, whereas all the hosts in the control group had high viremia levels. Despite minimal, if any, viremia, all six hosts generated neutralizing antibody titers close to those of the control monkeys while MV-directed cellular immunity reached levels at least as high as in wild-type-infected monkeys. These findings prove formally that efficient SLAM recognition is necessary for MV virulence and pathogenesis. They also suggest that the selectively SLAM-blind wild-type MV can be developed into a vaccine vector.
Figures

References
-
- Cattaneo, R., and M. B. Mc Chesney. 2008. Measles virus, p. 285-291. In B. Mahy and M. Van Regenmortel (ed.), In Encyclopedia of virology, 3rd ed., vol. 3. Academic Press, San Diego, CA.
-
- Cocks, B. G., C. C. Chang, J. M. Carballido, H. Yssel, J. E. de Vries, and G. Aversa. 1995. A novel receptor involved in T-cell activation. Nature 376:260-263. - PubMed
-
- Condack, C., J. C. Grivel, P. Devaux, L. Margolis, and R. Cattaneo. 2007. Measles virus vaccine attenuation: suboptimal infection of lymphatic tissue and tropism alteration. J. Infect. Dis. 196:541-549. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
