

Viral protein determinants of Lassa virus entry and release from polarized epithelial cells
- PMID: 20071570
- PMCID: PMC2838109
- DOI: 10.1128/JVI.02240-09
Viral protein determinants of Lassa virus entry and release from polarized epithelial cells
Abstract
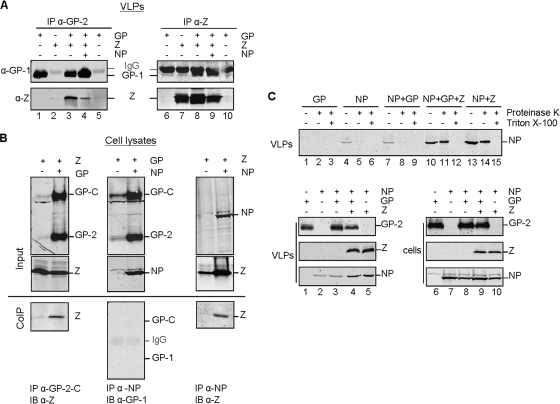
The epithelium plays a key role in the spread of Lassa virus. Transmission from rodents to humans occurs mainly via inhalation or ingestion of droplets, dust, or food contaminated with rodent urine. Here, we investigated Lassa virus infection in cultured epithelial cells and subsequent release of progeny viruses. We show that Lassa virus enters polarized Madin-Darby canine kidney (MDCK) cells mainly via the basolateral route, consistent with the basolateral localization of the cellular Lassa virus receptor alpha-dystroglycan. In contrast, progeny virus was efficiently released from the apical cell surface. Further, we determined the roles of the glycoprotein, matrix protein, and nucleoprotein in directed release of nascent virus. To do this, a virus-like-particle assay was developed in polarized MDCK cells based on the finding that, when expressed individually, both the glycoprotein GP and matrix protein Z form virus-like particles. We show that GP determines the apical release of Lassa virus from epithelial cells, presumably by recruiting the matrix protein Z to the site of virus assembly, which is in turn essential for nucleocapsid incorporation into virions.
Figures
Similar articles
-
Lassa virus Z protein is a matrix protein and sufficient for the release of virus-like particles [corrected].J Virol. 2003 Oct;77(19):10700-5. doi: 10.1128/jvi.77.19.10700-10705.2003. J Virol. 2003. PMID: 12970458 Free PMC article.
-
Inhibition of Lassa virus glycoprotein cleavage and multicycle replication by site 1 protease-adapted alpha(1)-antitrypsin variants.PLoS Negl Trop Dis. 2009 Jun 2;3(6):e446. doi: 10.1371/journal.pntd.0000446. PLoS Negl Trop Dis. 2009. PMID: 19488405 Free PMC article.
-
Basolateral budding of Marburg virus: VP40 retargets viral glycoprotein GP to the basolateral surface.J Infect Dis. 2007 Nov 15;196 Suppl 2:S232-6. doi: 10.1086/520584. J Infect Dis. 2007. PMID: 17940954
-
Lassa virus.Crit Rev Clin Lab Sci. 2004;41(4):339-90. doi: 10.1080/10408360490497456. Crit Rev Clin Lab Sci. 2004. PMID: 15487592 Review.
-
Vectorial Release of Human RNA Viruses from Epithelial Cells.Viruses. 2022 Jan 25;14(2):231. doi: 10.3390/v14020231. Viruses. 2022. PMID: 35215825 Free PMC article. Review.
Cited by
-
A role for the C terminus of Mopeia virus nucleoprotein in its incorporation into Z protein-induced virus-like particles.J Virol. 2010 May;84(10):5415-22. doi: 10.1128/JVI.02417-09. Epub 2010 Mar 3. J Virol. 2010. PMID: 20200234 Free PMC article.
-
Comprehensive Interactome Analysis Reveals that STT3B Is Required for N-Glycosylation of Lassa Virus Glycoprotein.J Virol. 2019 Nov 13;93(23):e01443-19. doi: 10.1128/JVI.01443-19. Print 2019 Dec 1. J Virol. 2019. PMID: 31511384 Free PMC article.
-
Identification of two functional domains within the arenavirus nucleoprotein.J Virol. 2011 Mar;85(5):2012-23. doi: 10.1128/JVI.01875-10. Epub 2010 Dec 15. J Virol. 2011. PMID: 21159858 Free PMC article.
-
Analysis of the Cell Type-Dependence on the Arenavirus Z-Mediated Virus-Like Particle Production.Front Microbiol. 2020 Sep 30;11:562814. doi: 10.3389/fmicb.2020.562814. eCollection 2020. Front Microbiol. 2020. PMID: 33117310 Free PMC article.
-
Human LAMP1 accelerates Lassa virus fusion and potently promotes fusion pore dilation upon forcing viral fusion with non-endosomal membrane.PLoS Pathog. 2022 Aug 15;18(8):e1010625. doi: 10.1371/journal.ppat.1010625. eCollection 2022 Aug. PLoS Pathog. 2022. PMID: 35969633 Free PMC article.
References
-
- Baize, S., J. Kaplon, C. Faure, D. Pannetier, M. C. Georges-Courbot, and V. Deubel. 2004. Lassa virus infection of human dendritic cells and macrophages is productive but fails to activate cells. J. Immunol. 172:2861-2869. - PubMed
-
- Ball, J. M., M. J. Mulligan, and R. W. Compans. 1997. Basolateral sorting of the HIV type 2 and SIV envelope glycoproteins in polarized epithelial cells: role of the cytoplasmic domain. AIDS Res. Hum. Retroviruses 13:665-675. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
