

Oxidative stress triggers the preferential assembly of base excision repair complexes on open chromatin regions
- PMID: 20071746
- PMCID: PMC2875005
- DOI: 10.1093/nar/gkp1247
Oxidative stress triggers the preferential assembly of base excision repair complexes on open chromatin regions
Abstract
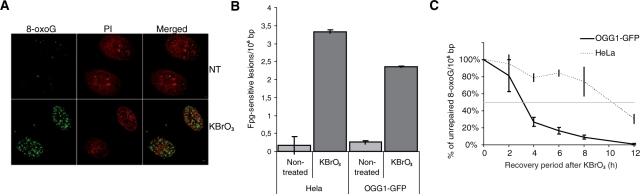
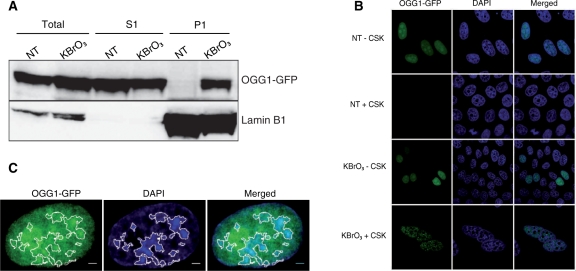
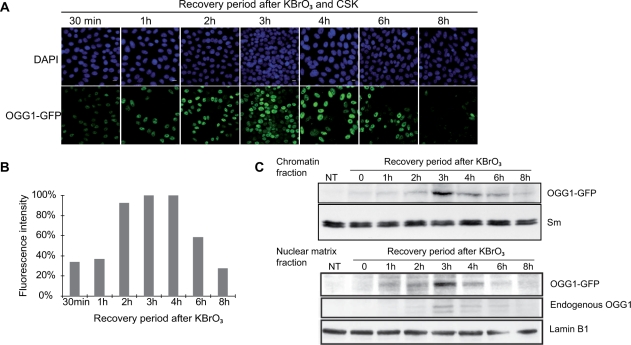
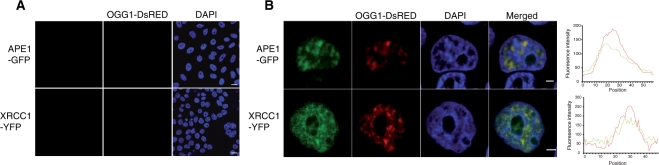
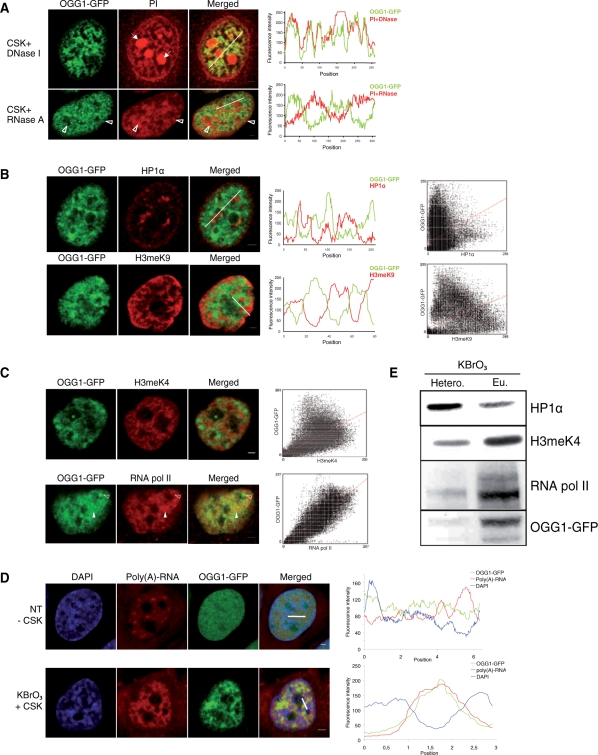
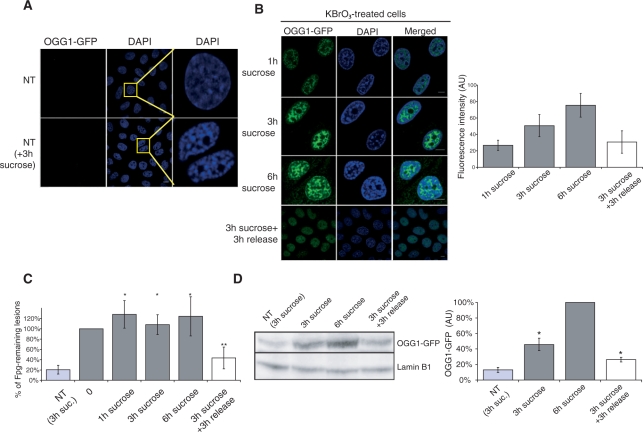
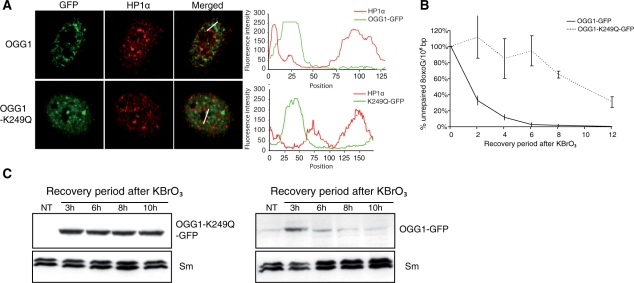
How DNA repair machineries detect and access, within the context of chromatin, lesions inducing little or no distortion of the DNA structure is a poorly understood process. Removal of oxidized bases is initiated by a DNA glycosylase that recognises and excises the damaged base, initiating the base excision repair (BER) pathway. We show that upon induction of 8-oxoguanine, a mutagenic product of guanine oxidation, the mammalian 8-oxoguanine DNA glycosylase OGG1 is recruited together with other proteins involved in BER to euchromatin regions rich in RNA and RNA polymerase II and completely excluded from heterochromatin. The underlying mechanism does not require direct interaction of the protein with the oxidized base, however, the release of the protein from the chromatin fraction requires completion of repair. Inducing chromatin compaction by sucrose results in a complete but reversible inhibition of the in vivo repair of 8-oxoguanine. We conclude that after induction of oxidative DNA damage, the DNA glycosylase is actively recruited to regions of open chromatin allowing the access of the BER machinery to the lesions, suggesting preferential repair of active chromosome regions.
Figures
References
-
- Barnes DE, Lindahl T. Repair and genetic consequences of endogenous DNA base damage in mammalian cells. Annu. Rev. Genet. 2004;38:445–476. - PubMed
-
- Shibutani S, Takeshita M, Grollman AP. Insertion of specific bases during DNA synthesis past the oxidation- damaged base 8-oxodG. Nature. 1991;349:431–434. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
