

Development and function of myeloid-derived suppressor cells generated from mouse embryonic and hematopoietic stem cells
- PMID: 20073041
- PMCID: PMC4370270
- DOI: 10.1002/stem.301
Development and function of myeloid-derived suppressor cells generated from mouse embryonic and hematopoietic stem cells
Abstract
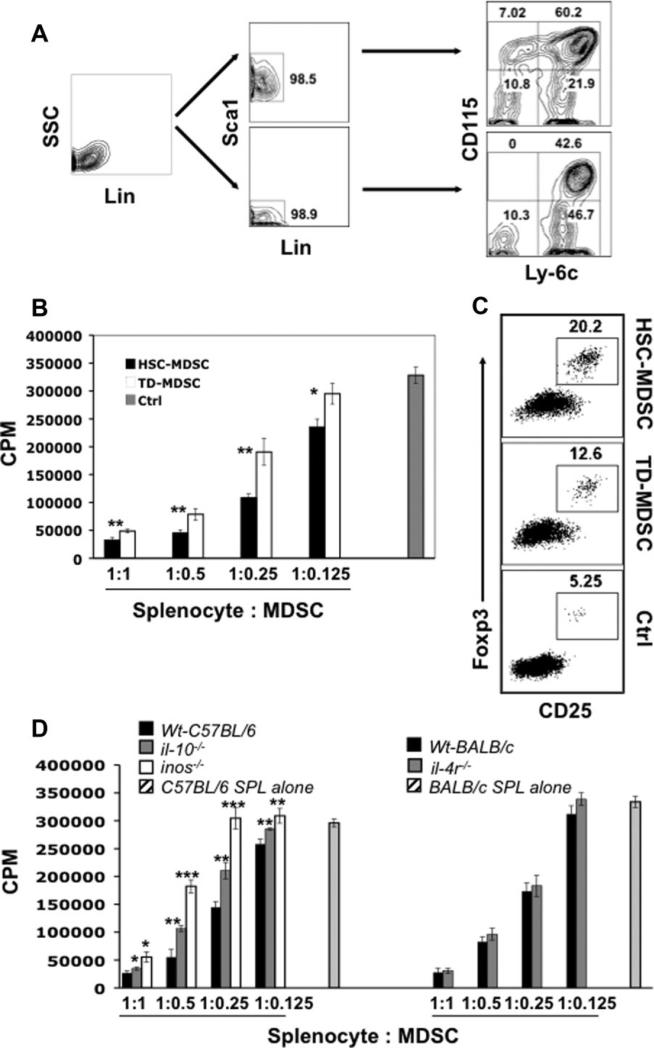
Emerging evidence suggests that myeloid-derived suppressor cells (MDSCs) have great potential as a novel immune intervention modality in the fields of transplantation and autoimmune diseases. Thus far, efforts to develop MDSC-based therapeutic strategies have been hampered by the lack of a reliable source of MDSCs. Here we show that functional MDSCs can be efficiently generated from mouse embryonic stem (ES) cells and bone marrow hematopoietic stem (HS) cells. In vitro-derived MDSCs encompass two homogenous subpopulations: CD115(+)Ly-6C(+) and CD115(+)Ly-6C(-) cells. The CD115(+)Ly-6C(+) subset is equivalent to the monocytic Gr-1(+)CD115(+)F4/80(+) MDSCs found in tumor-bearing mice. In contrast, the CD115(+)Ly-6C(-) cells, a previously unreported population of MDSCs, resemble the granulocyte/macrophage progenitors developmentally. In vitro, ES- and HS-MDSCs exhibit robust suppression against T-cell proliferation induced by polyclonal stimuli or alloantigens via multiple mechanisms involving nitric oxide synthase-mediated NO production and interleukin (IL)-10. Impressively, they display even stronger suppressive activity and significantly enhance ability to induce CD4(+)CD25(+)Foxp3(+) regulatory T-cell development compared with tumor-derived MDSCs. Furthermore, adoptive transfer of ES-MDSCs can effectively prevent alloreactive T-cell-mediated lethal graft-versus-host disease, leading to nearly 82% long-term survival among treated mice. The successful in vitro generation of MDSCs may represent a critical step toward potential clinical application of MDSCs.
Figures






References
-
- Serafini P, Borrello I, Bronte V. Myeloid suppressor cells in cancer: Recruitment, phenotype, properties, and mechanisms of immune suppression. Semin Cancer Biol. 2006;16:53–65. - PubMed
-
- Huang B, Pan PY, Li Q, et al. Gr-1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006;66:1123–1131. - PubMed
-
- Kusmartsev SA, Li Y, Chen SH. Gr-1+ myeloid cells derived from tumor-bearing mice inhibit primary T cell activation induced through CD3/CD28 costimulation. J Immunol. 2000;165:779–785. - PubMed
-
- Rodriguez PC, Quiceno DG, Zabaleta J, et al. Arginase I production in the tumor microenvironment by mature myeloid cells inhibits T-cell receptor expression and antigen-specific T-cell responses. Cancer Res. 2004;64:5839–5849. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
