

Temperature increase prevails over acidification in gene expression modulation of amastigote differentiation in Leishmania infantum
- PMID: 20074347
- PMCID: PMC2845110
- DOI: 10.1186/1471-2164-11-31
Temperature increase prevails over acidification in gene expression modulation of amastigote differentiation in Leishmania infantum
Abstract
Background: The extracellular promastigote and the intracellular amastigote stages alternate in the digenetic life cycle of the trypanosomatid parasite Leishmania. Amastigotes develop inside parasitophorous vacuoles of mammalian phagocytes, where they tolerate extreme environmental conditions. Temperature increase and pH decrease are crucial factors in the multifactorial differentiation process of promastigotes to amastigotes. Although expression profiling approaches for axenic, cell culture- and lesion-derived amastigotes have already been reported, the specific influence of temperature increase and acidification of the environment on developmental regulation of genes has not been previously studied. For the first time, we have used custom L. infantum genomic DNA microarrays to compare the isolated and the combined effects of both factors on the transcriptome.
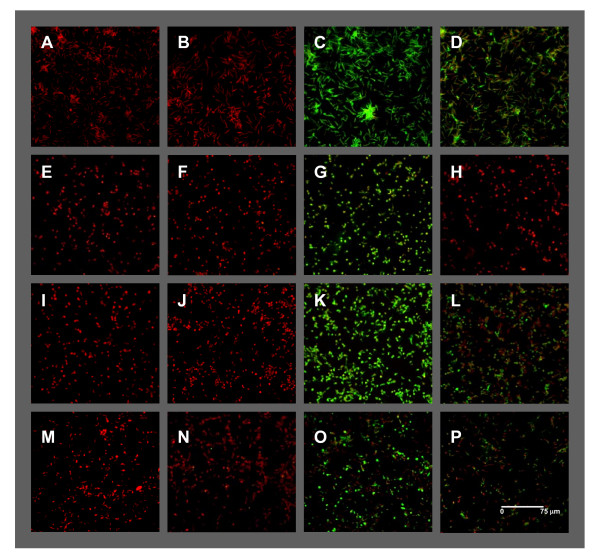
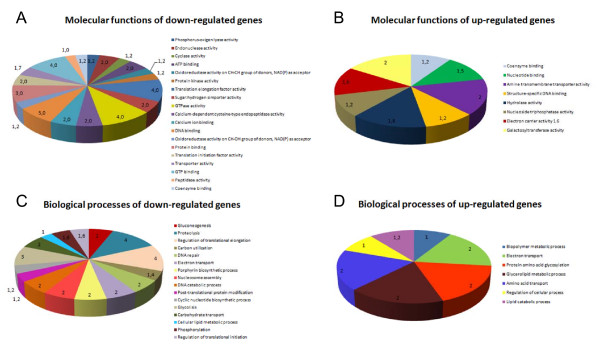
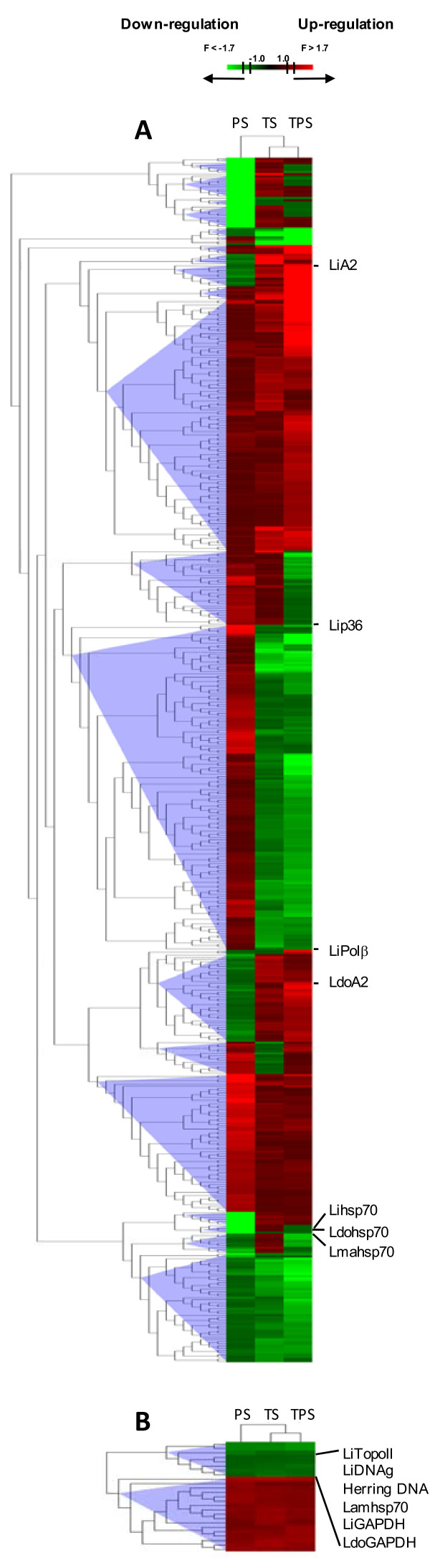
Results: Immunofluorescence analysis of promastigote-specific glycoprotein gp46 and expression modulation analysis of the amastigote-specific A2 gene have revealed that concomitant exposure to temperature increase and acidification leads to amastigote-like forms. The temperature-induced gene expression profile in the absence of pH variation resembles the profile obtained under combined exposure to both factors unlike that obtained for exposure to acidification alone. In fact, the subsequent fold change-based global iterative hierarchical clustering analysis supports these findings.
Conclusions: The specific influence of temperature and pH on the differential regulation of genes described in this study and the evidence provided by clustering analysis is consistent with the predominant role of temperature increase over extracellular pH decrease in the amastigote differentiation process, which provides new insights into Leishmania physiology.
Figures

References
-
- Pral EM, da Moitinho ML, Balanco JM, Teixeira VR, Milder RV, Alfieri SC. Growth phase and medium ph modulate the expression of proteinase activities and the development of megasomes in axenically cultivated Leishmania (Leishmania) amazonensis amastigote-like organisms. J Parasitol. 2003;89(1):35–43. doi: 10.1645/0022-3395(2003)089[0035:GPAMPM]2.0.CO;2. - DOI - PubMed
