

Sarcolemmal ATP-sensitive K(+) channels control energy expenditure determining body weight
- PMID: 20074528
- PMCID: PMC2849280
- DOI: 10.1016/j.cmet.2009.11.009
Sarcolemmal ATP-sensitive K(+) channels control energy expenditure determining body weight
Abstract
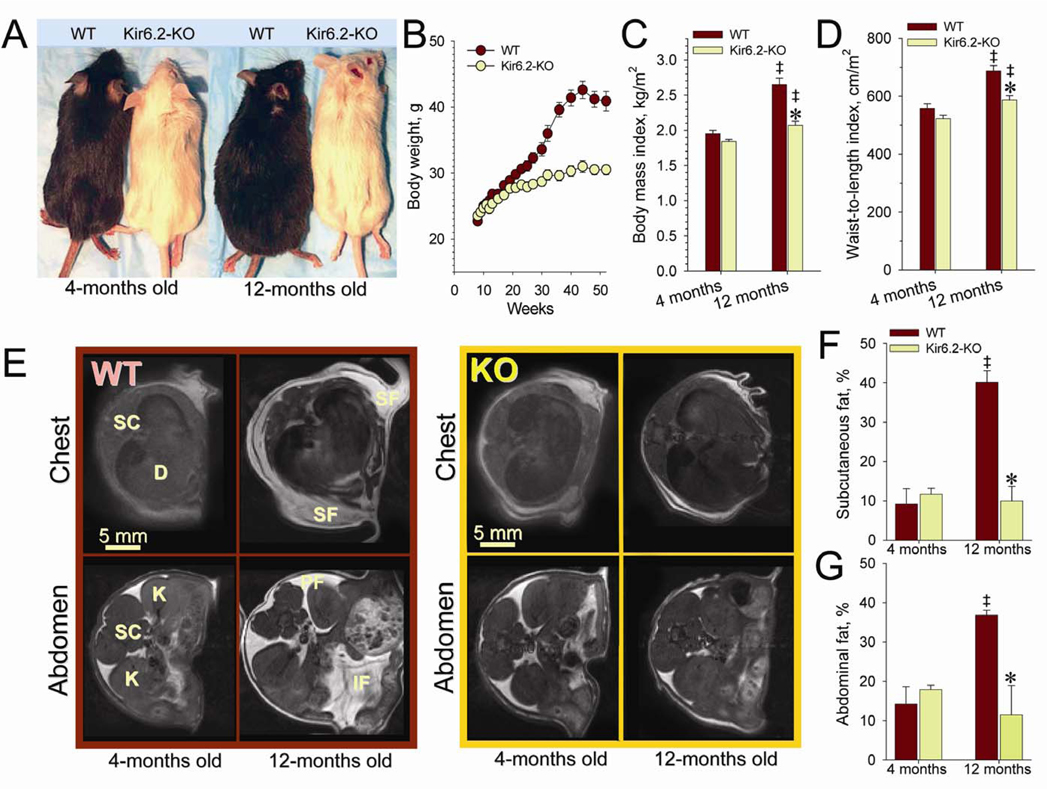
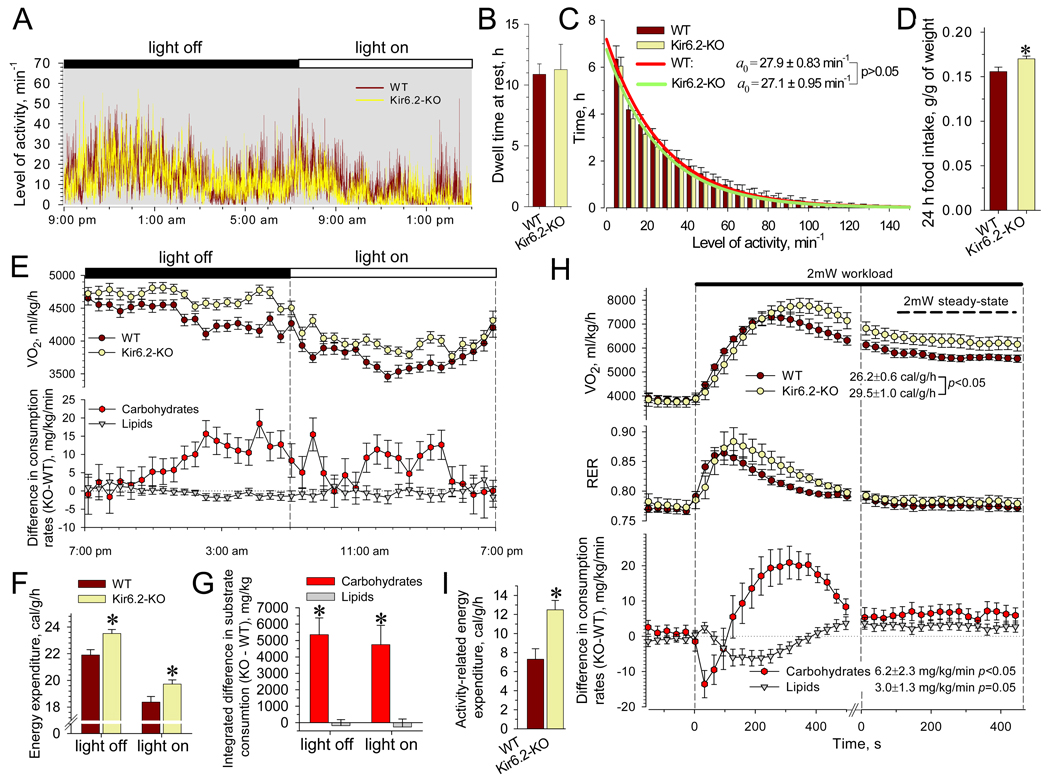
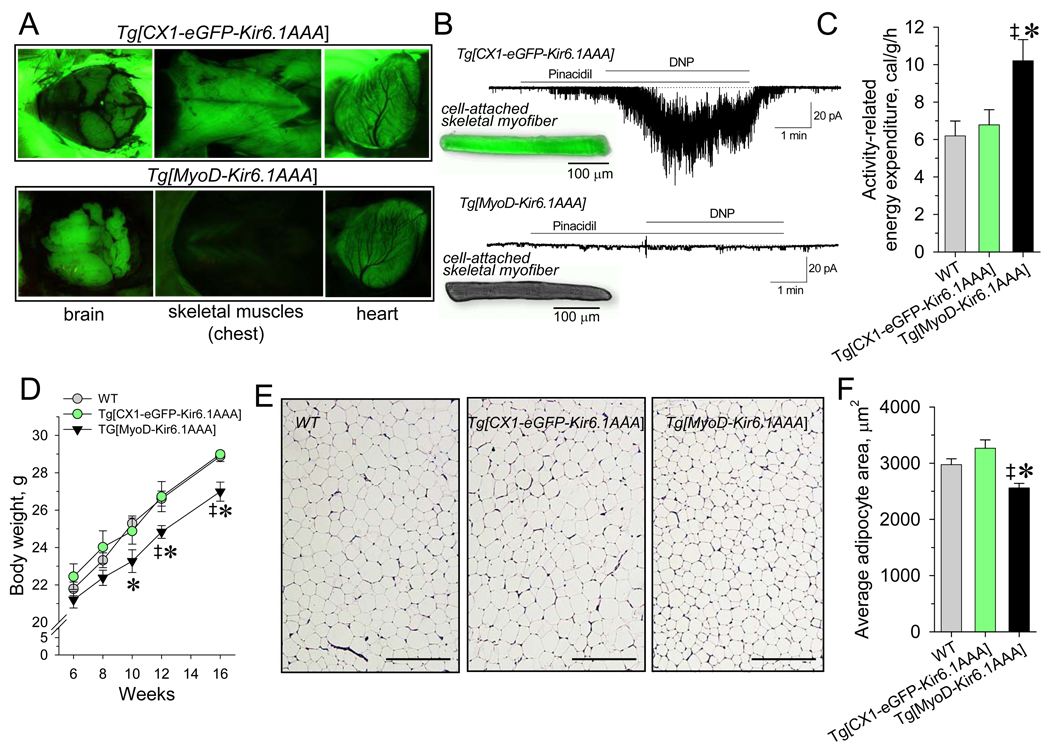
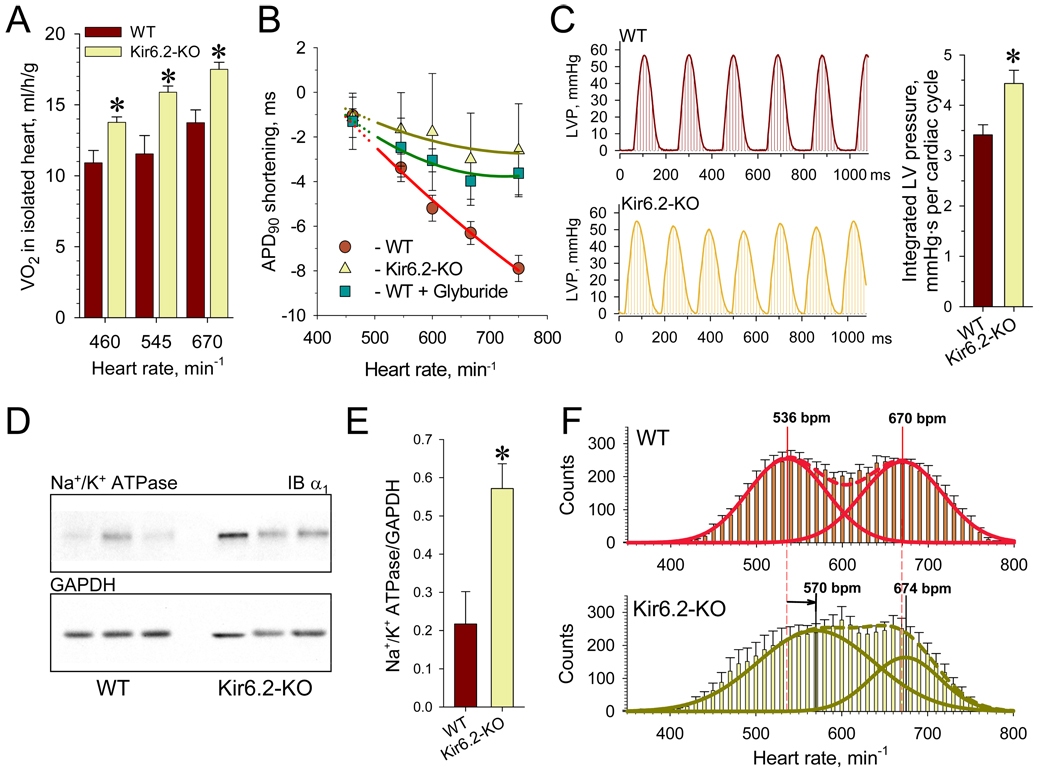
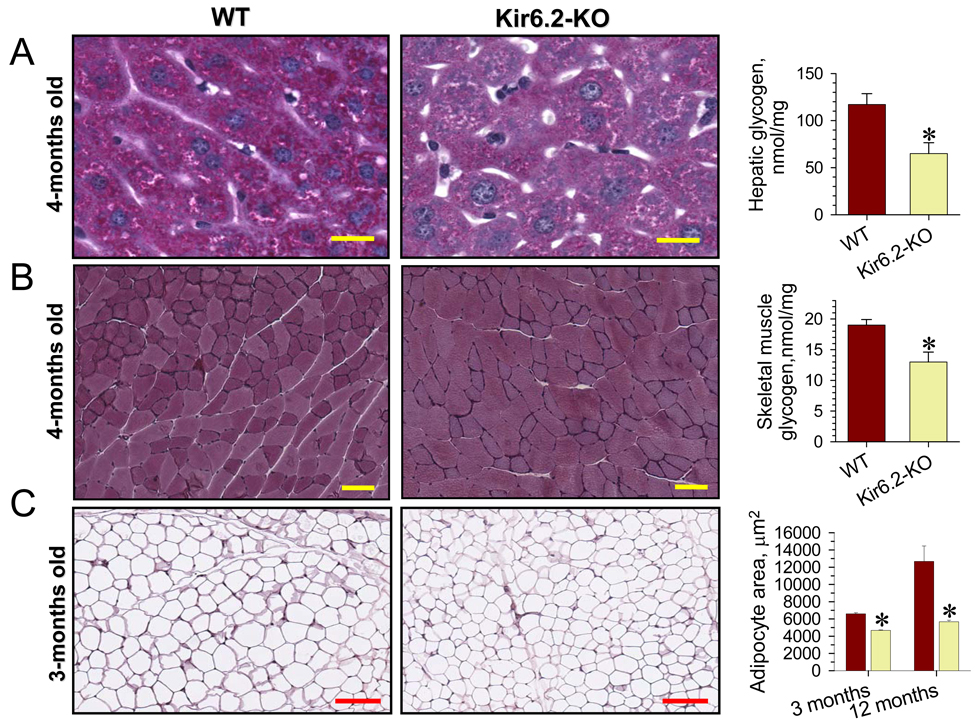
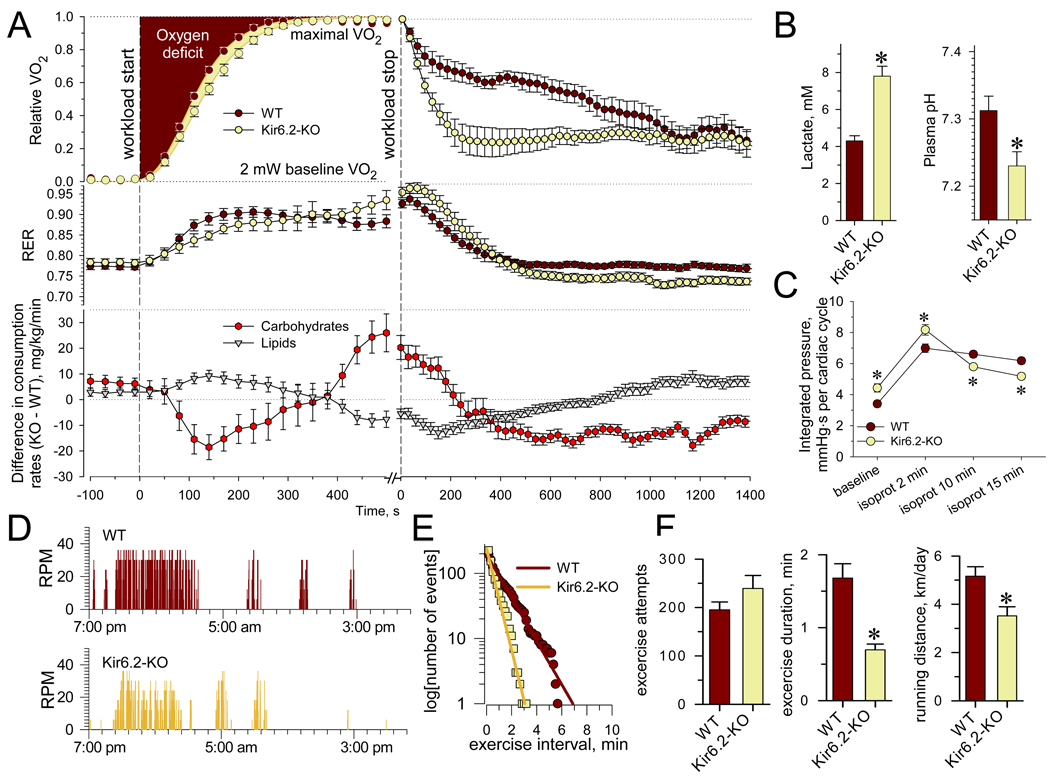
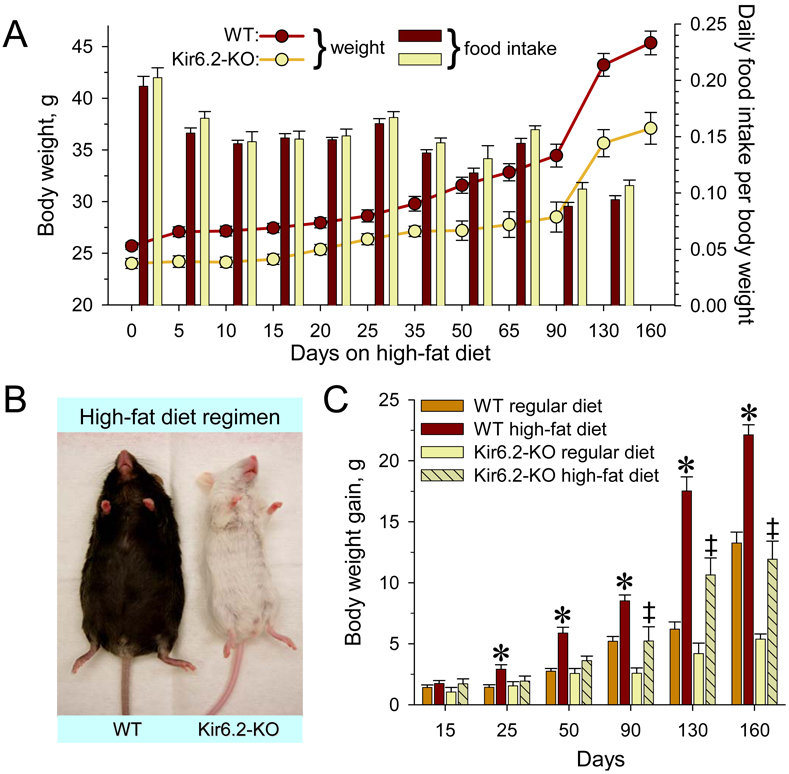
Metabolic processes that regulate muscle energy use are major determinants of bodily energy balance. Here, we find that sarcolemmal ATP-sensitive K(+) (K(ATP)) channels, which couple membrane excitability with cellular metabolic pathways, set muscle energy expenditure under physiological stimuli. Disruption of K(ATP) channel function provoked, under conditions of unaltered locomotor activity and blood substrate availability, an extra energy cost of cardiac and skeletal muscle performance. Inefficient fuel metabolism in K(ATP) channel-deficient striated muscles reduced glycogen and fat body depots, promoting a lean phenotype. The propensity to lesser body weight imposed by K(ATP) channel deficit persisted under a high-fat diet, yet obesity restriction was achieved at the cost of compromised physical endurance. Thus, sarcolemmal K(ATP) channels govern muscle energy economy, and their downregulation in a tissue-specific manner could present an antiobesity strategy by rendering muscle increasingly thermogenic at rest and less fuel efficient during exercise.
2010 Elsevier Inc.
Figures
References
-
- Abraham MR, Selivanov VA, Hodgson D, Pucar D, Zingman LV, Wieringa B, Dzeja PP, Alekseev AE, Terzic A. Coupling of cell energetics with membrane metabolic sensing: Integrative signaling through creatine kinase phosphor-transfer disrupted by M-CK gene knockout. J. Biol. Chem. 2002;277:24427–24434. - PubMed
-
- Aguilar-Bryan L, Bryan J, Nakazaki M. Of mice and men: KATP channels and insulin secretion. Recent Prog. Horm. Res. 2001;56:47–68. - PubMed
-
- Babenko AP, Aguilar-Bryan L, Bryan J. A view of SUR/KIR6.X, KATP channels. Annu. Rev. Physiol. 1998;60:667–687. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
