

Studying Cerebellar Circuits by Remote Control of Selected Neuronal Types with GABA(A) Receptors
- PMID: 20076763
- PMCID: PMC2805427
- DOI: 10.3389/neuro.02.029.2009
Studying Cerebellar Circuits by Remote Control of Selected Neuronal Types with GABA(A) Receptors
Abstract
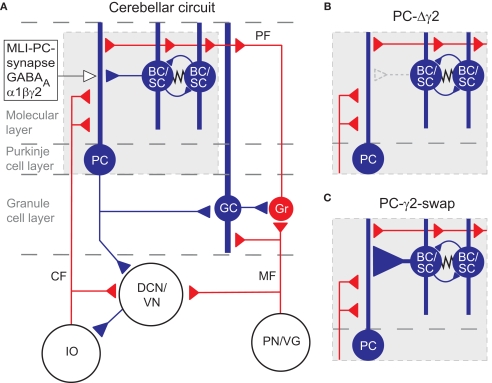
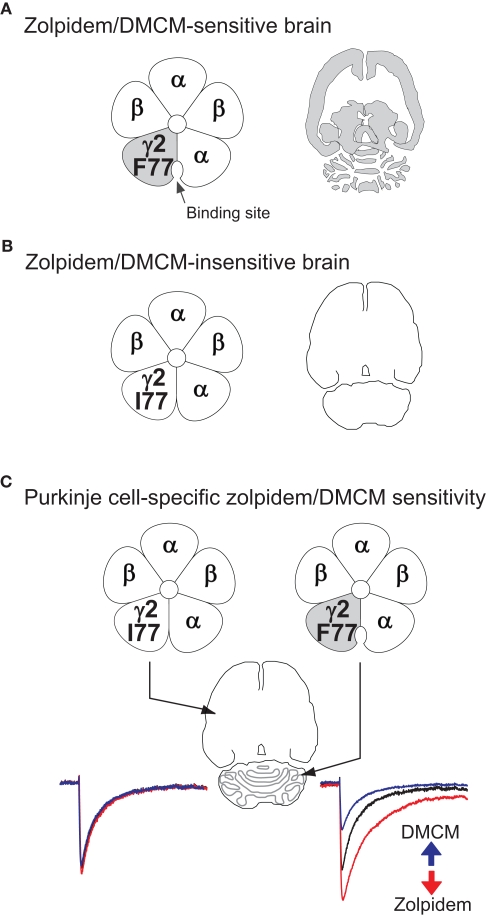
Although GABA(A) receptor-mediated inhibition of cerebellar Purkinje cells by molecular layer interneurons (MLIs) has been studied intensely at the cellular level, it has remained unclear how this inhibition regulates cerebellum-dependent behaviour. We have implemented two complementary approaches to investigate the function of the MLI-Purkinje cell synapse on the behavioural level. In the first approach we permanently disrupted inhibitory fast synaptic transmission at the synapse by genetically removing the postsynaptic GABA(A) receptors from Purkinje cells (PC-Deltagamma2 mice). We found that chronic disruption of the MLI-Purkinje cell synapse strongly impaired cerebellar learning of the vestibular occular reflex (VOR), presumably by disrupting the temporal patterns of Purkinje cell activity. However, in PC-Deltagamma2 mice the baseline VOR reflex was only mildly affected; indeed PC-Deltagamma2 mice show no ataxia or gait abnormalities, suggesting that MLI control of Purkinje cell activity is either not involved in ongoing motor tasks or that the system compensates for its loss. To investigate the latter possibility we developed an alternative genetic technique; we made the MLI-Purkinje cell synapse selectively sensitive to rapid manipulation with the GABA(A) receptor modulator zolpidem (PC-gamma2-swap mice). Minutes after intraperitoneal zolpidem injection, these PC-gamma2-swap mice developed severe motor abnormalities, revealing a substantial contribution of the MLI-Purkinje cell synapses to real time motor control. The cell-type selective permanent knockout of synaptic GABAergic input and the fast reversible modulation of GABAergic input at the same synapse illustrate how pursuing both strategies gives a fuller view.
Keywords: memory consolidation; purkinje cell; zolpidem; β-carboline; γ-aminobutyric acid type A receptor.
Figures
Similar articles
-
Interneuronal NMDA receptors regulate long-term depression and motor learning in the cerebellum.J Physiol. 2019 Feb;597(3):903-920. doi: 10.1113/JP276794. Epub 2018 Nov 24. J Physiol. 2019. PMID: 30382582 Free PMC article.
-
Expression of a Form of Cerebellar Motor Memory Requires Learned Alterations to the Activity of Inhibitory Molecular Layer Interneurons.J Neurosci. 2023 Jan 25;43(4):601-612. doi: 10.1523/JNEUROSCI.0731-22.2022. Epub 2022 Dec 9. J Neurosci. 2023. PMID: 36639897 Free PMC article.
-
Facial stimulation induces long-term depression at cerebellar molecular layer interneuron-Purkinje cell synapses in vivo in mice.Front Cell Neurosci. 2015 Jun 9;9:214. doi: 10.3389/fncel.2015.00214. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26106296 Free PMC article.
-
Modulation, Plasticity and Pathophysiology of the Parallel Fiber-Purkinje Cell Synapse.Front Synaptic Neurosci. 2016 Nov 3;8:35. doi: 10.3389/fnsyn.2016.00035. eCollection 2016. Front Synaptic Neurosci. 2016. PMID: 27857688 Free PMC article. Review.
-
Molecular Layer Interneurons: Key Elements of Cerebellar Network Computation and Behavior.Neuroscience. 2021 May 10;462:22-35. doi: 10.1016/j.neuroscience.2020.10.008. Epub 2020 Oct 17. Neuroscience. 2021. PMID: 33075461 Review.
Cited by
-
Removal of GABA(A) receptor γ2 subunits from parvalbumin neurons causes wide-ranging behavioral alterations.PLoS One. 2011;6(9):e24159. doi: 10.1371/journal.pone.0024159. Epub 2011 Sep 2. PLoS One. 2011. PMID: 21912668 Free PMC article.
-
Kit Ligand and Kit receptor tyrosine kinase sustain synaptic inhibition of Purkinje cells.Elife. 2024 Mar 27;12:RP89792. doi: 10.7554/eLife.89792. Elife. 2024. PMID: 38536959 Free PMC article.
-
Cerebellar disease associated with anti-glutamic acid decarboxylase antibodies: review.J Neural Transm (Vienna). 2017 Oct;124(10):1171-1182. doi: 10.1007/s00702-017-1754-3. Epub 2017 Jul 8. J Neural Transm (Vienna). 2017. PMID: 28689294 Review.
-
Sepsis Impairs Purkinje Cell Functions and Motor Behaviors Through Microglia Activation.Cerebellum. 2024 Apr;23(2):329-339. doi: 10.1007/s12311-023-01531-7. Epub 2023 Feb 15. Cerebellum. 2024. PMID: 36790600
-
Chemical genetics: receptor-ligand pairs for rapid manipulation of neuronal activity.Curr Opin Neurobiol. 2012 Feb;22(1):54-60. doi: 10.1016/j.conb.2011.10.008. Epub 2011 Nov 24. Curr Opin Neurobiol. 2012. PMID: 22119143 Free PMC article. Review.
References
-
- Albus J. S. (1971). A theory of cerebellar function. Math. Biosci 10, 25–6110.1016/0025-5564(71)90051-4 - DOI
-
- Alexander G. M., Rogan S. C., Abbas A. I., Armbruster B. N., Pei Y., Allen J. A., Nonneman R. J., Hartmann J., Moy S. S., Nicolelis M. A., McNamara J. O., Roth B. L. (2009). Remote control of neuronal activity in transgenic mice expressing evolved G protein-coupled receptors. Neuron 63, 27–3910.1016/j.neuron.2009.06.014 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources