

The pathogenicity determinant of Citrus tristeza virus causing the seedling yellows syndrome maps at the 3'-terminal region of the viral genome
- PMID: 20078776
- PMCID: PMC6640426
- DOI: 10.1111/j.1364-3703.2009.00572.x
The pathogenicity determinant of Citrus tristeza virus causing the seedling yellows syndrome maps at the 3'-terminal region of the viral genome
Abstract
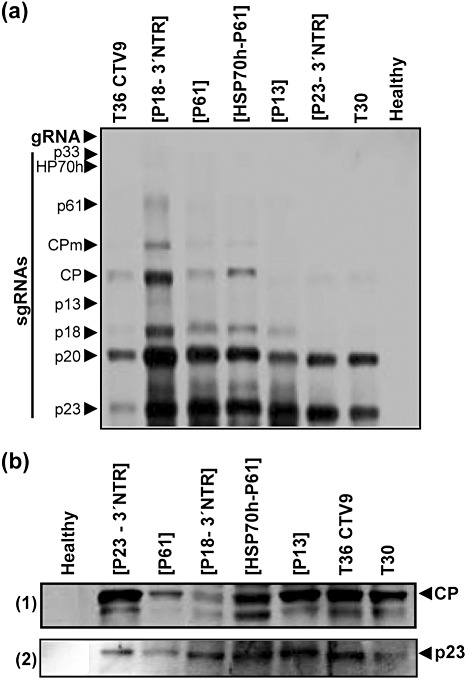
Citrus tristeza virus (CTV) (genus Closterovirus, family Closteroviridae) causes some of the more important viral diseases of citrus worldwide. The ability to map disease-inducing determinants of CTV is needed to develop better diagnostic and disease control procedures. A distinctive phenotype of some isolates of CTV is the ability to induce seedling yellows (SY) in sour orange, lemon and grapefruit seedlings. In Florida, the decline isolate of CTV, T36, induces SY, whereas a widely distributed mild isolate, T30, does not. To delimit the viral sequences associated with the SY syndrome, we created a number of T36/T30 hybrids by substituting T30 sequences into different regions of the 3' half of the genome of an infectious cDNA of T36. Eleven T36/T30 hybrids replicated in Nicotiana benthamiana protoplasts. Five of these hybrids formed viable virions that were mechanically transmitted to Citrus macrophylla, a permissive host for CTV. All induced systemic infections, similar to that of the parental T36 clone. Tissues from these C. macrophylla source plants were then used to graft inoculate sour orange and grapefruit seedlings. Inoculation with three of the T30/T36 hybrid constructs induced SY symptoms identical to those of T36; however, two hybrids with T30 substitutions in the p23-3' nontranslated region (NTR) (nucleotides 18 394-19 296) failed to induce SY. Sour orange seedlings infected with a recombinant non-SY p23-3' NTR hybrid also remained symptomless when challenged with the parental virus (T36), demonstrating the potential feasibility of using engineered constructs of CTV to mitigate disease.
Figures


References
-
- Albiach‐Martí, M.R. , Guerri, J. , Cambra, M. , Garnsey, S.M. and Moreno, P. (2000a) Differentiation of citrus tristeza virus isolates by serological analysis of the p25 coat protein peptide maps. J. Virol. Methods, 88, 25–34. - PubMed
-
- Albiach‐Martí, M.R. , Guerri, J. , Hermoso de Mendoza, A. , Laigret, F. , Ballester‐Olmos, J.F. and Moreno, P. (2000b) Aphid transmission alters the genomic and defective RNA populations of citrus tristeza virus. Phytopathology, 90, 134–138. - PubMed
-
- Albiach‐Martí, M.R. , Mawassi, M. , Gowda, S. , Satyanarayana, T. , Hilf, M.E. , Shanker, S. , Almira, E.C. , Vives, M.C. , López, C. , Guerri, J. , Flores, R. , Moreno, P. , Garnsey, S.M. and Dawson, W.O. (2000c) Sequences of Citrus tristeza virus separated in time and space are essentially identical. J. Virol. 74, 6856–6865. - PMC - PubMed
-
- Bar‐Joseph, M. and Dawson, W.O. (2008) Citrus tristeza virus In: Encyclopedia of Virology, Vol. 1 (Mahy B.W.J. and Van Regenmortel M.H.V., eds), pp. 520–525. Oxford: Elsevier Ltd.
-
- Bar‐Joseph, M. , Marcus, R. and Lee, R.F. (1989) The continuous challenge of citrus tristeza virus control. Annu. Rev. Phytopathol. 27, 291–316.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
