

Molecular mechanisms of polyploidy and hybrid vigor
- PMID: 20080432
- PMCID: PMC2821985
- DOI: 10.1016/j.tplants.2009.12.003
Molecular mechanisms of polyploidy and hybrid vigor
Abstract
Hybrids such as maize (Zea mays) or domestic dog (Canis lupus familiaris) grow bigger and stronger than their parents. This is also true for allopolyploids such as wheat (Triticum spp.) or frog (i.e. Xenopus and Silurana) that contain two or more sets of chromosomes from different species. The phenomenon, known as hybrid vigor or heterosis, was systematically characterized by Charles Darwin (1876). The rediscovery of heterosis in maize a century ago has revolutionized plant and animal breeding and production. Although genetic models for heterosis have been rigorously tested, the molecular bases remain elusive. Recent studies have determined the roles of nonadditive gene expression, small RNAs, and epigenetic regulation, including circadian-mediated metabolic pathways, in hybrid vigor, which could lead to better use and exploitation of the increased biomass and yield in hybrids and allopolyploids for food, feed, and biofuels.
Copyright 2010 Elsevier Ltd. All rights reserved.
Figures
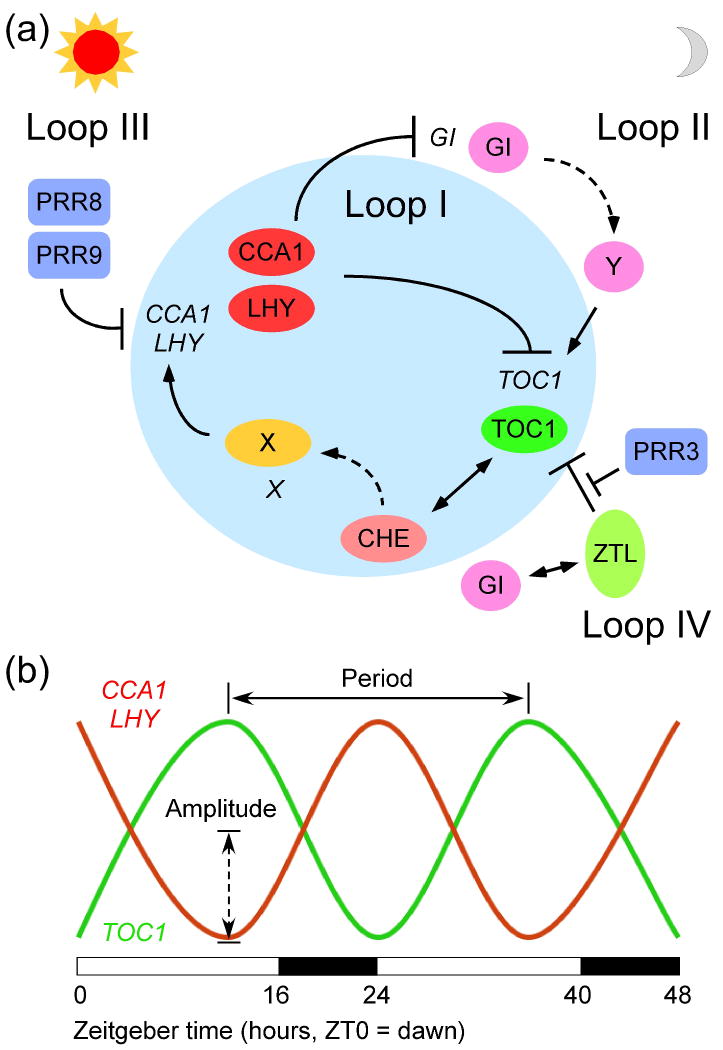

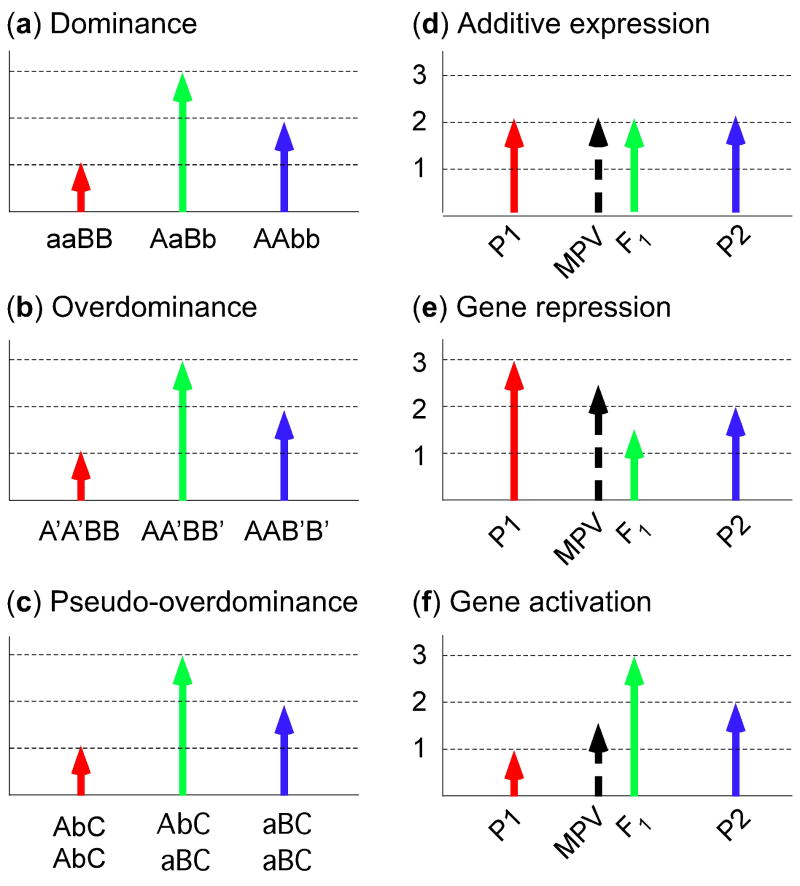
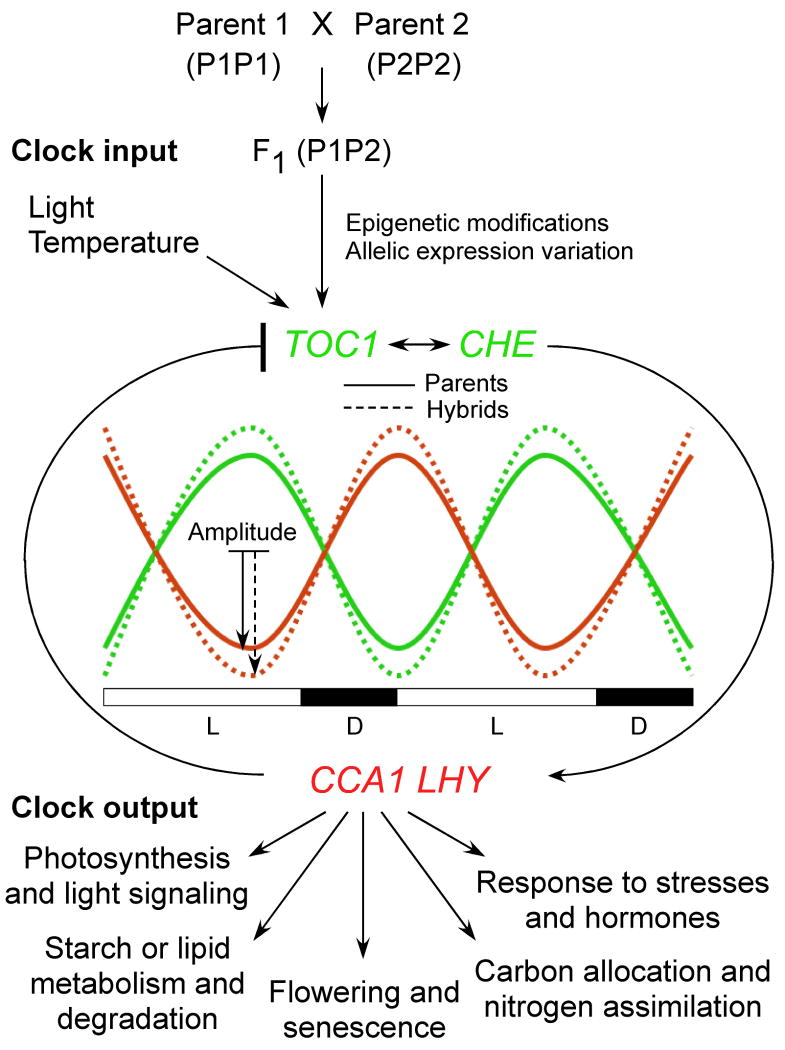
References
-
- Darwin CR. The Effects of Cross- and Self-fertilization in the Vegetable Kingdom. John Murry; 1876.
-
- Shull GH. The composition of a field of maize. Amer Breeders Assoc Rep. 1908;4:296–301.
-
- East EM. Reports of the Connecticut Agricultural Experiment Station for Years 1907-1908. Connecticut Agricultural Experiment Station; 1908. Inbreeding in corn; pp. 419–428.
-
- Duvick DN. Biotechnology in the 1930s: the development of hybrid maize. Nat Rev Genet. 2001;2:69–74. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
