

Ras membrane orientation and nanodomain localization generate isoform diversity
- PMID: 20080631
- PMCID: PMC2824305
- DOI: 10.1073/pnas.0903907107
Ras membrane orientation and nanodomain localization generate isoform diversity
Abstract
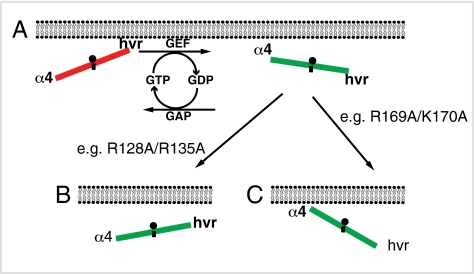
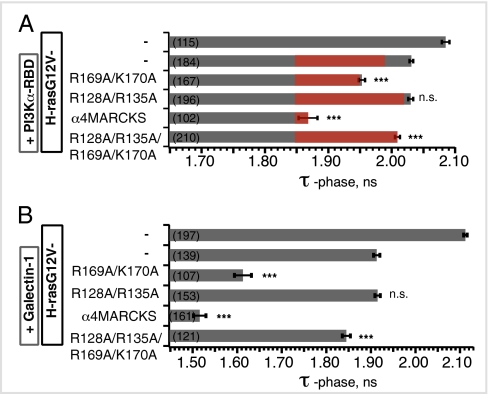
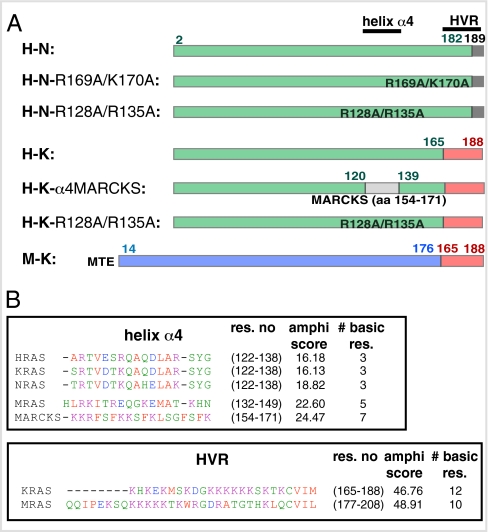
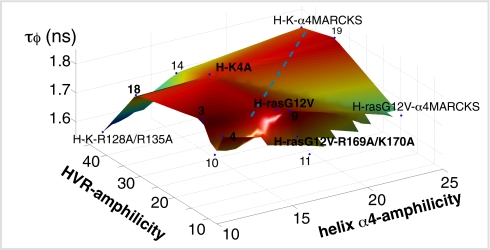
The structural elements encoding functional diversity among Ras GTPases are poorly defined. The orientation of the G domain of H-ras with respect to the plane of the plasma membrane is recognized by the Ras binding domain of C-Raf, coupling orientation to MAPK activation. We now show that two other proteins, phosphoinositide-3-kinase-alpha and the structurally unrelated galectin-1, also recognize G-domain orientation. These results rationalize the role of galectin-1 in generating active GTP-H-ras signaling nanoclusters. However, molecular dynamics simulations of K-ras membrane insertion and fluorescence lifetime imaging microscopy (FLIM)-Förster resonance energy transfer (FRET) imaging of the effector interactions of N-Ras, K-Ras, and M-ras suggest that there are two hyperactive, signaling-competent orientations of the Ras G domain. Mutational and functional analyses establish a clear relationship between effector binding and the amphilicities of helix alpha4 and the C-terminal hypervariable region, thus confirming that these structural elements critically tune the orientation of the Ras G domain. Finally, we show that G-domain orientation and nanoclustering synergize to generate Ras isoform specificity with respect to effector interactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
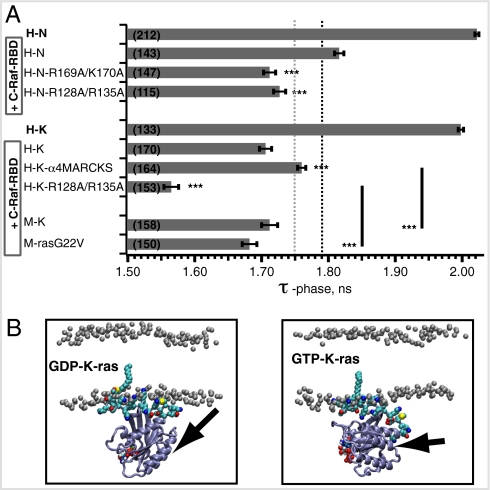
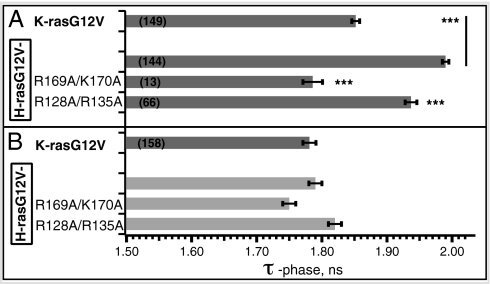
References
-
- Wennerberg K, Rossman KL, Der CJ. The Ras superfamily at a glance. J Cell Sci. 2005;118:843–846. - PubMed
-
- Vetter IR, Wittinghofer A. The guanine nucleotide-binding switch in three dimensions. Science. 2001;294:1299–1304. - PubMed
-
- Sprang SR. G protein mechanisms: Insights from structural analysis. Annu Rev Biochem. 1997;66:639–678. - PubMed
-
- Herrmann C. Ras-effector interactions: After one decade. Curr Opin Struct Biol. 2003;13:122–129. - PubMed
-
- Herrmann C, Nassar N. Ras and its effectors. Prog Biophys Mol Biol. 1996;66:1–41. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
