

Anthrax toxin triggers the activation of src-like kinases to mediate its own uptake
- PMID: 20080640
- PMCID: PMC2824395
- DOI: 10.1073/pnas.0910782107
Anthrax toxin triggers the activation of src-like kinases to mediate its own uptake
Abstract
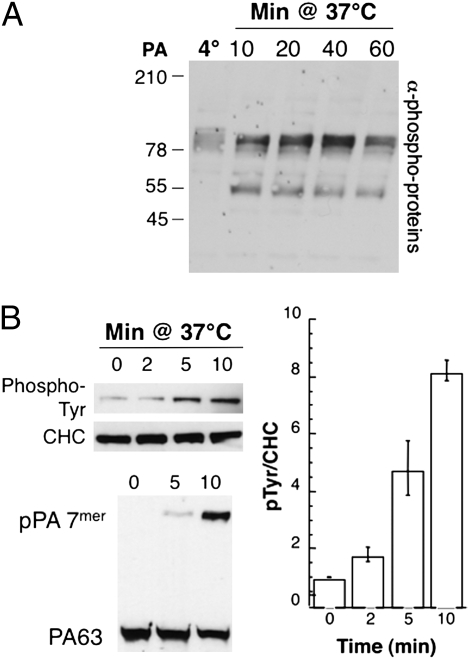
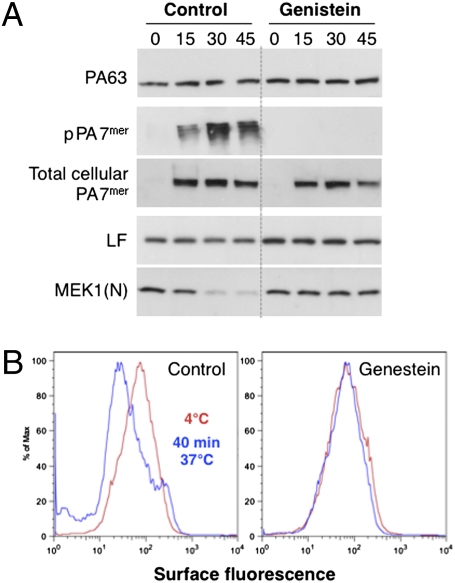
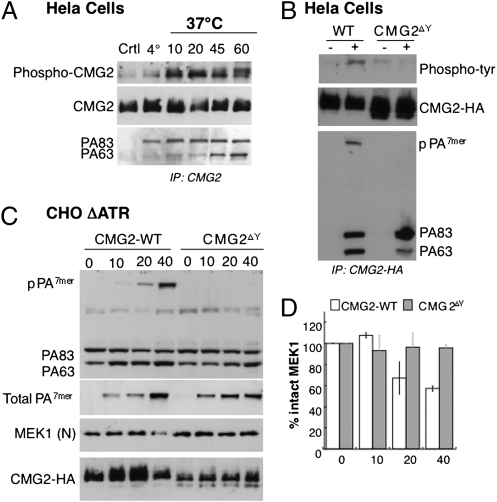
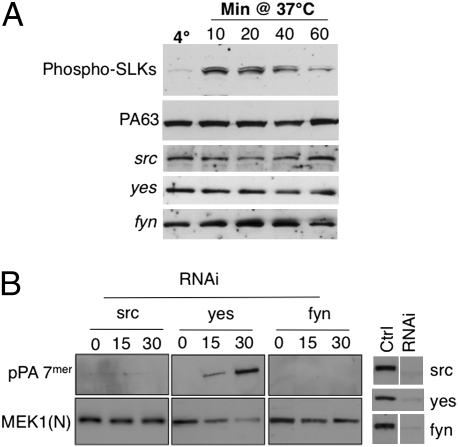
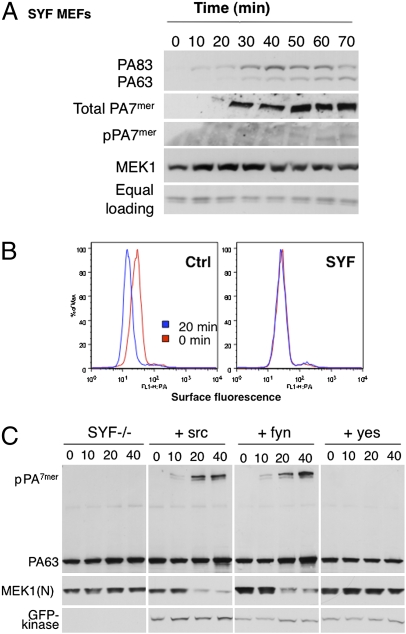
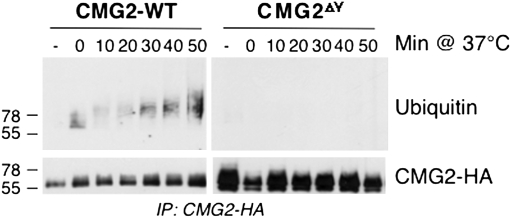
AB-type toxins, like other bacterial toxins, are notably opportunistic molecules. They rely on target cell receptors to reach the appropriate location within the target cell where translocation of their enzymatic subunits occurs. The anthrax toxin, however, times its own uptake, suggesting that toxin binding triggers specific signaling events. Here we show that the anthrax toxin triggers tyrosine phosphorylation of its own receptors, capillary morphogenesis gene 2 and tumor endothelial marker 8, which are not endowed with intrinsic kinase activity. This is required for efficient toxin uptake because endocytosis of the mutant receptor lacking the cytoplasmic tyrosine residues is strongly delayed. Phosphorylation of the receptors was dependent on src-like kinases, which where activated upon toxin binding. Importantly, src-dependent phosphorylation of the receptor was required for its subsequent ubiquitination, which in turn was required for clathrin-mediated endocytosis. Consistently, we found that uptake of the anthrax toxin and processing of the lethal factor substrate MEK1 are inhibited by silencing of src and fyn, as well as in src and fyn knockout cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Endocytosis of the anthrax toxin is mediated by clathrin, actin and unconventional adaptors.PLoS Pathog. 2010 Mar 5;6(3):e1000792. doi: 10.1371/journal.ppat.1000792. PLoS Pathog. 2010. PMID: 20221438 Free PMC article.
-
Anthrax toxin triggers endocytosis of its receptor via a lipid raft-mediated clathrin-dependent process.J Cell Biol. 2003 Feb 3;160(3):321-8. doi: 10.1083/jcb.200211018. Epub 2003 Jan 27. J Cell Biol. 2003. PMID: 12551953 Free PMC article.
-
Functional interactions between anthrax toxin receptors and the WNT signalling protein LRP6.Cell Microbiol. 2008 Dec;10(12):2509-19. doi: 10.1111/j.1462-5822.2008.01226.x. Epub 2008 Aug 20. Cell Microbiol. 2008. PMID: 18717822
-
Interactions between anthrax toxin receptors and protective antigen.Curr Opin Microbiol. 2005 Feb;8(1):106-12. doi: 10.1016/j.mib.2004.12.005. Curr Opin Microbiol. 2005. PMID: 15694864 Review.
-
Roles of Anthrax Toxin Receptor 2 in Anthrax Toxin Membrane Insertion and Pore Formation.Toxins (Basel). 2016 Jan 22;8(2):34. doi: 10.3390/toxins8020034. Toxins (Basel). 2016. PMID: 26805886 Free PMC article. Review.
Cited by
-
Bacillus anthracis Protective Antigen Shows High Specificity for a UV Induced Mouse Model of Cutaneous Squamous Cell Carcinoma.Front Med (Lausanne). 2019 Feb 12;6:22. doi: 10.3389/fmed.2019.00022. eCollection 2019. Front Med (Lausanne). 2019. PMID: 30809524 Free PMC article.
-
The Ins and Outs of Anthrax Toxin.Toxins (Basel). 2016 Mar 10;8(3):69. doi: 10.3390/toxins8030069. Toxins (Basel). 2016. PMID: 26978402 Free PMC article. Review.
-
Protein Lipidation by Palmitate Controls Macrophage Function.Cells. 2022 Feb 6;11(3):565. doi: 10.3390/cells11030565. Cells. 2022. PMID: 35159374 Free PMC article. Review.
-
Endocytosis of the anthrax toxin is mediated by clathrin, actin and unconventional adaptors.PLoS Pathog. 2010 Mar 5;6(3):e1000792. doi: 10.1371/journal.ppat.1000792. PLoS Pathog. 2010. PMID: 20221438 Free PMC article.
-
Hyaline fibromatosis syndrome with mutation c.1074delT of the CMG2 gene: a case report.J Med Case Rep. 2014 Sep 3;8:291. doi: 10.1186/1752-1947-8-291. J Med Case Rep. 2014. PMID: 25186005 Free PMC article.
References
-
- Schiavo G, van der Goot FG. The bacterial toxin toolkit. Nat Rev Mol Cell Biol. 2001;2:530–537. - PubMed
-
- Sandvig K, van deurs B. Delivery into cells: Lessons learned from plant and bacterial toxins. Gene Ther. 2005;12:865–872. - PubMed
-
- Sandvig K, van Deurs B. Transport of protein toxins into cells: Pathways used by ricin, cholera toxin and Shiga toxin. FEBS Lett. 2002;529:49–53. - PubMed
-
- Gruenberg J, van der Goot FG. Mechanisms of pathogen entry through the endosomal compartments. Nat Rev Mol Cell Biol. 2006;7:495–504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
