

Stk40 links the pluripotency factor Oct4 to the Erk/MAPK pathway and controls extraembryonic endoderm differentiation
- PMID: 20080709
- PMCID: PMC2824367
- DOI: 10.1073/pnas.0905657107
Stk40 links the pluripotency factor Oct4 to the Erk/MAPK pathway and controls extraembryonic endoderm differentiation
Abstract
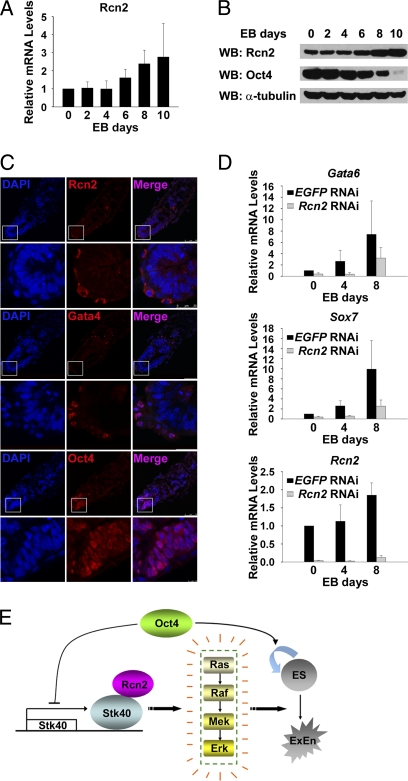
Self-renewal and differentiation of embryonic stem cells (ESCs) are controlled by intracellular transcriptional factors and extracellular factor-activated signaling pathways. Transcription factor Oct4 is a key player maintaining ESCs in an undifferentiated state, whereas the Erk/MAPK pathway is known to be important for ESC differentiation. However, the manner in which intracellular pluripotency factors modulate extracellular factor-activated signaling pathways in ESCs is not well understood. Here, we report identification of a target gene of Oct4, serine/threonine kinase 40 (Stk40), which is able to activate the Erk/MAPK pathway and induce extraembryonic-endoderm (ExEn) differentiation in mouse ESCs. Interestingly, cells overexpressing Stk40 exclusively contribute to the ExEn layer of chimeric embryos when injected into host blastocysts. In contrast, deletion of Stk40 in ESCs markedly reduces ExEn differentiation in vitro. Mechanistically, Stk40 interacts with Rcn2, which also activates Erk1/2 to induce ExEn specification in mouse ESCs. Moreover, Rcn2 proteins are specifically located in the cytoplasm of the ExEn layer of early mouse embryos. Importantly, knockdown of Rcn2 blocks Stk40-activated Erk1/2 and ESC differentiation. Therefore, our study establishes a link between the pluripotency factor Oct4 and the Erk/MAPK signaling pathway, and it uncovers cooperating signals in the Erk/MAPK activation that control ExEn differentiation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Deletion of STK40 protein in mice causes respiratory failure and death at birth.J Biol Chem. 2013 Feb 22;288(8):5342-52. doi: 10.1074/jbc.M112.409433. Epub 2013 Jan 4. J Biol Chem. 2013. PMID: 23293024 Free PMC article.
-
Dual functions of T-box 3 (Tbx3) in the control of self-renewal and extraembryonic endoderm differentiation in mouse embryonic stem cells.J Biol Chem. 2011 Mar 11;286(10):8425-8436. doi: 10.1074/jbc.M110.202150. Epub 2010 Dec 28. J Biol Chem. 2011. PMID: 21189255 Free PMC article.
-
Isolation of Oct4-expressing extraembryonic endoderm precursor cell lines.PLoS One. 2009 Sep 28;4(9):e7216. doi: 10.1371/journal.pone.0007216. PLoS One. 2009. PMID: 19784378 Free PMC article.
-
Pelota regulates the development of extraembryonic endoderm through activation of bone morphogenetic protein (BMP) signaling.Stem Cell Res. 2014 Jul;13(1):61-74. doi: 10.1016/j.scr.2014.04.011. Epub 2014 Apr 26. Stem Cell Res. 2014. PMID: 24835669
-
MAPK/ERK Pathway as a Central Regulator in Vertebrate Organ Regeneration.Int J Mol Sci. 2022 Jan 27;23(3):1464. doi: 10.3390/ijms23031464. Int J Mol Sci. 2022. PMID: 35163418 Free PMC article. Review.
Cited by
-
Single-stranded DNA binding protein Ssbp3 induces differentiation of mouse embryonic stem cells into trophoblast-like cells.Stem Cell Res Ther. 2016 May 28;7(1):79. doi: 10.1186/s13287-016-0340-1. Stem Cell Res Ther. 2016. PMID: 27236334 Free PMC article.
-
High STK40 Expression as an Independent Prognostic Biomarker and Correlated with Immune Infiltrates in Low-Grade Gliomas.Int J Gen Med. 2021 Oct 5;14:6389-6400. doi: 10.2147/IJGM.S335821. eCollection 2021. Int J Gen Med. 2021. PMID: 34675607 Free PMC article.
-
Characterization of Ath29, a major mouse atherosclerosis susceptibility locus, and identification of Rcn2 as a novel regulator of cytokine expression.Am J Physiol Heart Circ Physiol. 2011 Sep;301(3):H1056-61. doi: 10.1152/ajpheart.00366.2011. Epub 2011 Jun 10. Am J Physiol Heart Circ Physiol. 2011. PMID: 21666121 Free PMC article.
-
Deletion of STK40 protein in mice causes respiratory failure and death at birth.J Biol Chem. 2013 Feb 22;288(8):5342-52. doi: 10.1074/jbc.M112.409433. Epub 2013 Jan 4. J Biol Chem. 2013. PMID: 23293024 Free PMC article.
-
Stem cells and calcium signaling.Adv Exp Med Biol. 2012;740:891-916. doi: 10.1007/978-94-007-2888-2_40. Adv Exp Med Biol. 2012. PMID: 22453975 Free PMC article. Review.
References
-
- Rossant J. Stem cells and lineage development in the mammalian blastocyst. Reprod Fertil Dev. 2007;19:111–118. - PubMed
-
- Tam PP, Loebel DA. Gene function in mouse embryogenesis: get set for gastrulation. Nat Rev Genet. 2007;8:368–381. - PubMed
-
- Bielinska M, Narita N, Wilson DB. Distinct roles for visceral endoderm during embryonic mouse development. Int J Dev Biol. 1999;43:183–205. - PubMed
-
- Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292:154–156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
