

Structural basis for cofactor-independent dioxygenation of N-heteroaromatic compounds at the alpha/beta-hydrolase fold
- PMID: 20080731
- PMCID: PMC2818892
- DOI: 10.1073/pnas.0909033107
Structural basis for cofactor-independent dioxygenation of N-heteroaromatic compounds at the alpha/beta-hydrolase fold
Abstract
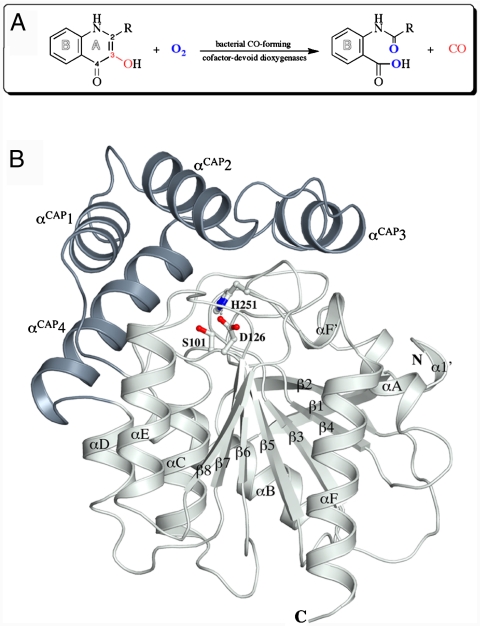
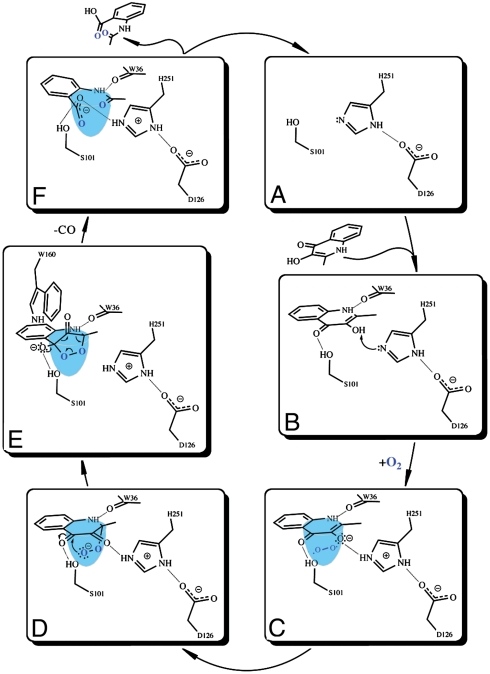
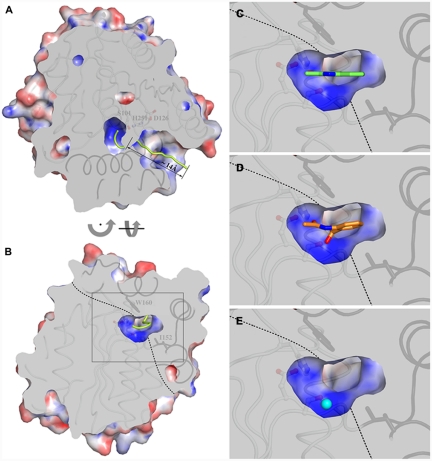
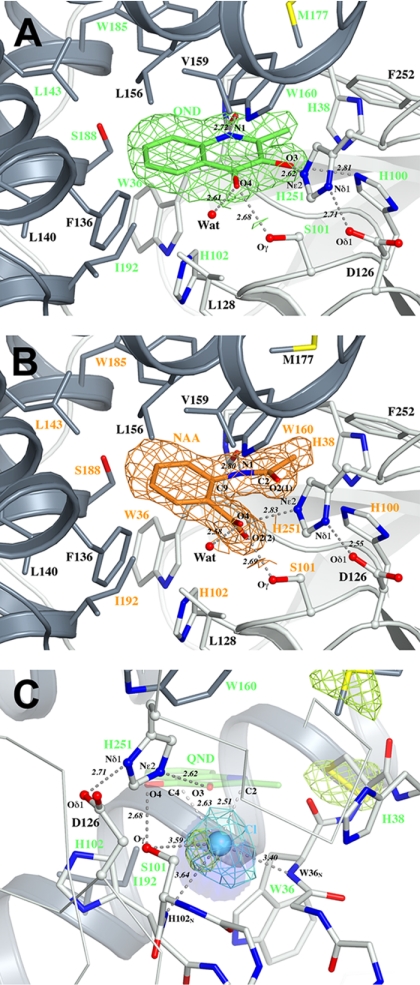
Enzymatic catalysis of oxygenation reactions in the absence of metal or organic cofactors is a considerable biochemical challenge. The CO-forming 1-H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase (HOD) from Arthrobacter nitroguajacolicus Rü61a and 1-H-3-hydroxy-4-oxoquinoline 2,4-dioxygenase (QDO) from Pseudomonas putida 33/1 are homologous cofactor-independent dioxygenases involved in the breakdown of N-heteroaromatic compounds. To date, they are the only dioxygenases suggested to belong to the alpha/beta-hydrolase fold superfamily. Members of this family typically catalyze hydrolytic processes rather than oxygenation reactions. We present here the crystal structures of both HOD and QDO in their native state as well as the structure of HOD in complex with its natural 1-H-3-hydroxy-4-oxoquinaldine substrate, its N-acetylanthranilate reaction product, and chloride as dioxygen mimic. HOD and QDO are structurally very similar. They possess a classical alpha/beta-hydrolase fold core domain additionally equipped with a cap domain. Organic substrates bind in a preorganized active site with an orientation ideally suited for selective deprotonation of their hydroxyl group by a His/Asp charge-relay system affording the generation of electron-donating species. The "oxyanion hole" of the alpha/beta-hydrolase fold, typically employed to stabilize the tetrahedral intermediate in ester hydrolysis reactions, is utilized here to host and control oxygen chemistry, which is proposed to involve a peroxide anion intermediate. Product release by proton back transfer from the catalytic histidine is driven by minimization of intramolecular charge repulsion. Structural and kinetic data suggest a nonnucleophilic general-base mechanism. Our analysis provides a framework to explain cofactor-independent dioxygenation within a protein architecture generally employed to catalyze hydrolytic reactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Bacterial 2,4-dioxygenases: new members of the alpha/beta hydrolase-fold superfamily of enzymes functionally related to serine hydrolases.J Bacteriol. 1999 Sep;181(18):5725-33. doi: 10.1128/JB.181.18.5725-5733.1999. J Bacteriol. 1999. PMID: 10482514 Free PMC article.
-
2,4-dioxygenases catalyzing N-heterocyclic-ring cleavage and formation of carbon monoxide. Purification and some properties of 1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase from Arthrobacter sp. Rü61a and comparison with 1H-3-hydroxy-4-oxoquinoline 2,4-dioxygenase from Pseudomonas putida 33/1.Eur J Biochem. 1996 Sep 15;240(3):576-83. doi: 10.1111/j.1432-1033.1996.0576h.x. Eur J Biochem. 1996. PMID: 8856057
-
Dioxygenases without requirement for cofactors and their chemical model reaction: compulsory order ternary complex mechanism of 1H-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase involving general base catalysis by histidine 251 and single-electron oxidation of the substrate dianion.Biochemistry. 2004 Nov 16;43(45):14485-99. doi: 10.1021/bi048735u. Biochemistry. 2004. PMID: 15533053
-
Ring-cleaving dioxygenases with a cupin fold.Appl Environ Microbiol. 2012 Apr;78(8):2505-14. doi: 10.1128/AEM.07651-11. Epub 2012 Jan 27. Appl Environ Microbiol. 2012. PMID: 22287012 Free PMC article. Review.
-
A Paradigm for CH Bond Cleavage: Structural and Functional Aspects of Transition State Stabilization by Mandelate Racemase.Adv Protein Chem Struct Biol. 2017;109:113-160. doi: 10.1016/bs.apcsb.2017.04.007. Epub 2017 Jun 9. Adv Protein Chem Struct Biol. 2017. PMID: 28683916 Review.
Cited by
-
Immunosuppressant mycophenolic acid biosynthesis employs a new globin-like enzyme for prenyl side chain cleavage.Acta Pharm Sin B. 2019 Nov;9(6):1253-1258. doi: 10.1016/j.apsb.2019.06.009. Epub 2019 Jun 28. Acta Pharm Sin B. 2019. PMID: 31867170 Free PMC article.
-
The Metal Drives the Chemistry: Dual Functions of Acireductone Dioxygenase.Chem Rev. 2017 Aug 9;117(15):10474-10501. doi: 10.1021/acs.chemrev.7b00117. Epub 2017 Jul 21. Chem Rev. 2017. PMID: 28731690 Free PMC article. Review.
-
The multiple signaling systems regulating virulence in Pseudomonas aeruginosa.Microbiol Mol Biol Rev. 2012 Mar;76(1):46-65. doi: 10.1128/MMBR.05007-11. Microbiol Mol Biol Rev. 2012. PMID: 22390972 Free PMC article. Review.
-
Molecular basis for cyclooxygenase inhibition by the non-steroidal anti-inflammatory drug naproxen.J Biol Chem. 2010 Nov 5;285(45):34950-9. doi: 10.1074/jbc.M110.162982. Epub 2010 Sep 1. J Biol Chem. 2010. PMID: 20810665 Free PMC article.
-
Origin of the proton-transfer step in the cofactor-free (1H)-3-hydroxy-4-oxoquinaldine 2,4-dioxygenase: effect of the basicity of an active site His residue.J Biol Chem. 2014 Mar 21;289(12):8620-32. doi: 10.1074/jbc.M113.543033. Epub 2014 Jan 30. J Biol Chem. 2014. PMID: 24482238 Free PMC article.
References
-
- Fetzner S. Oxygenases without requirement for cofactors or metal ions. Appl Microbiol Biotechnol. 2002;60:243–257. - PubMed
-
- Fetzner S. Cofactor-independent oxygenases go it alone. Nat Chem Biol. 2007;3:374–375. - PubMed
-
- Tseng CC, Vaillancourt FH, Bruner SD, Walsh CT. DpgC is a metal- and cofactor-free 3,5-dihydroxyphenylacetyl-CoA 1,2-dioxygenase in the vancomycin biosynthetic pathway. Chem Biol. 2004;11:1195–1203. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
