

Saccharomyces cerevisiae Msh2-Msh6 DNA binding kinetics reveal a mechanism of targeting sites for DNA mismatch repair
- PMID: 20080735
- PMCID: PMC2818965
- DOI: 10.1073/pnas.0908302107
Saccharomyces cerevisiae Msh2-Msh6 DNA binding kinetics reveal a mechanism of targeting sites for DNA mismatch repair
Abstract
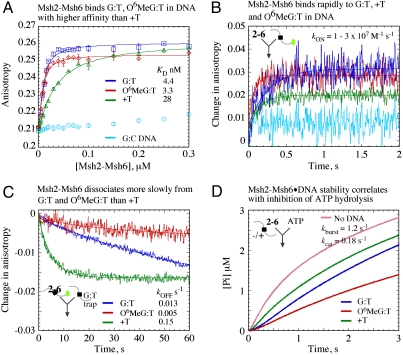
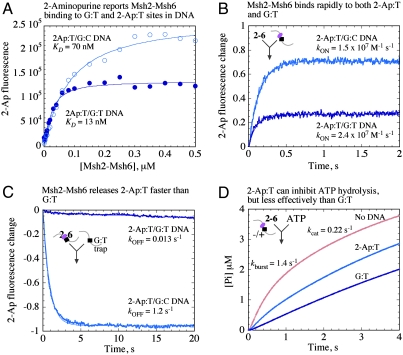
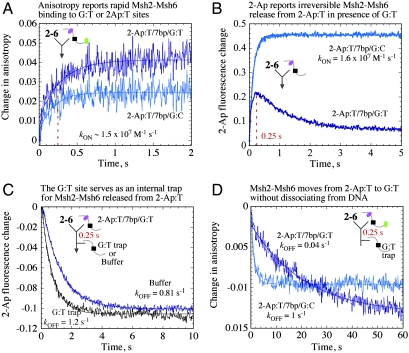
The DNA mismatch repair system (MMR) identifies replication errors and damaged bases in DNA and functions to preserve genomic integrity. MutS performs the task of locating mismatched base pairs, loops and lesions and initiating MMR, and the fundamental question of how this protein targets specific sites in DNA is unresolved. To address this question, we examined the interactions between Saccharomyces cerevisiae Msh2-Msh6, a eukaryotic MutS homolog, and DNA in real time. The reaction kinetics reveal that Msh2-Msh6 binds a variety of sites at similarly fast rates (k (ON) approximately 10(7) M(-1) s(-1)), and its selectivity manifests in differential dissociation rates; e.g., the protein releases a 2-Aminopurine:T base pair approximately 90-fold faster than a G:T mismatch. On releasing the 2-Ap:T site, Msh2-Msh6 is able to move laterally on DNA to locate a nearby G:T site. The long-lived Msh2-Msh6.G:T complex triggers the next step in MMR--formation of an ATP-bound clamp--more effectively than the short-lived Msh2-Msh6.2-Ap:T complex. Mutation of Glu in the conserved Phe-X-Glu DNA binding motif stabilizes Msh2-Msh6(E339A).2-Ap:T complex, and the mutant can signal 2-Ap:T repair as effectively as wild-type Msh2-Msh6 signals G:T repair. These findings suggest a targeting mechanism whereby Msh2-Msh6 scans DNA, interrogating base pairs by transient contacts and pausing at potential target sites, and the longer the pause the greater the likelihood of MMR.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Iyer RR, Pluciennik A, Burdett V, Modrich PL. DNA mismatch repair: Functions and mechanisms. Chem Rev. 2006;106(2):302–323. - PubMed
-
- Kunkel TA, Erie DA. DNA Mismatch Repair. Annu Rev Biochem. 2005;74:681–710. - PubMed
-
- Jiricny J. The multifaceted mismatch-repair system. Nat Rev Mol Cell Biol. 2006;7(5):335–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
