

Conformational transitions in DNA polymerase I revealed by single-molecule FRET
- PMID: 20080740
- PMCID: PMC2818957
- DOI: 10.1073/pnas.0910909107
Conformational transitions in DNA polymerase I revealed by single-molecule FRET
Abstract
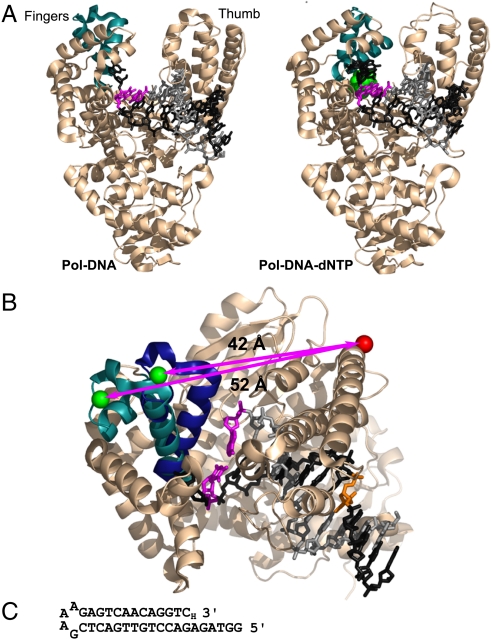
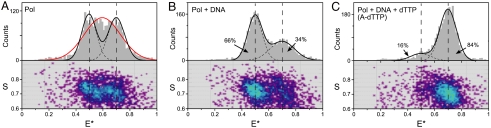
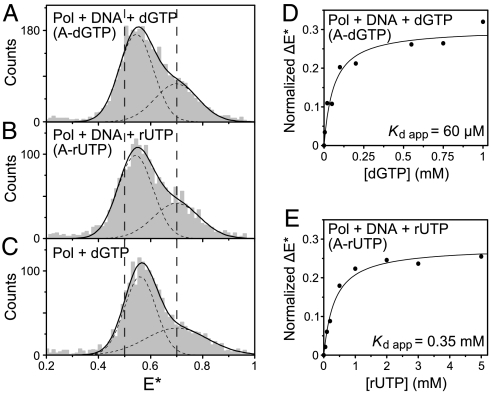
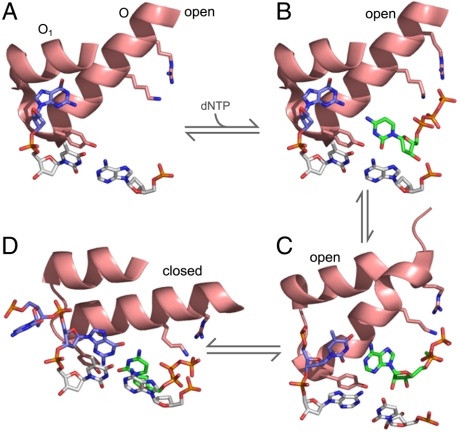
The remarkable fidelity of most DNA polymerases depends on a series of early steps in the reaction pathway which allow the selection of the correct nucleotide substrate, while excluding all incorrect ones, before the enzyme is committed to the chemical step of nucleotide incorporation. The conformational transitions that are involved in these early steps are detectable with a variety of fluorescence assays and include the fingers-closing transition that has been characterized in structural studies. Using DNA polymerase I (Klenow fragment) labeled with both donor and acceptor fluorophores, we have employed single-molecule fluorescence resonance energy transfer to study the polymerase conformational transitions that precede nucleotide addition. Our experiments clearly distinguish the open and closed conformations that predominate in Pol-DNA and Pol-DNA-dNTP complexes, respectively. By contrast, the unliganded polymerase shows a broad distribution of FRET values, indicating a high degree of conformational flexibility in the protein in the absence of its substrates; such flexibility was not anticipated on the basis of the available crystallographic structures. Real-time observation of conformational dynamics showed that most of the unliganded polymerase molecules sample the open and closed conformations in the millisecond timescale. Ternary complexes formed in the presence of mismatched dNTPs or complementary ribonucleotides show unique FRET species, which we suggest are relevant to kinetic checkpoints that discriminate against these incorrect substrates.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Conformational landscapes of DNA polymerase I and mutator derivatives establish fidelity checkpoints for nucleotide insertion.Nat Commun. 2013;4:2131. doi: 10.1038/ncomms3131. Nat Commun. 2013. PMID: 23831915 Free PMC article.
-
dNTP-dependent conformational transitions in the fingers subdomain of Klentaq1 DNA polymerase: insights into the role of the "nucleotide-binding" state.J Biol Chem. 2013 May 10;288(19):13575-91. doi: 10.1074/jbc.M112.432690. Epub 2013 Mar 22. J Biol Chem. 2013. PMID: 23525110 Free PMC article.
-
Distinct complexes of DNA polymerase I (Klenow fragment) for base and sugar discrimination during nucleotide substrate selection.J Biol Chem. 2011 Apr 22;286(16):14480-92. doi: 10.1074/jbc.M111.218750. Epub 2011 Feb 28. J Biol Chem. 2011. PMID: 21362617 Free PMC article.
-
DNA polymerase fidelity: kinetics, structure, and checkpoints.Biochemistry. 2004 Nov 16;43(45):14317-24. doi: 10.1021/bi048422z. Biochemistry. 2004. PMID: 15533035 Review.
-
Prokaryotic DNA polymerase I: evolution, structure, and "base flipping" mechanism for nucleotide selection.J Mol Biol. 2001 May 18;308(5):823-37. doi: 10.1006/jmbi.2001.4619. J Mol Biol. 2001. PMID: 11352575 Review.
Cited by
-
Protein Ensembles: How Does Nature Harness Thermodynamic Fluctuations for Life? The Diverse Functional Roles of Conformational Ensembles in the Cell.Chem Rev. 2016 Jun 8;116(11):6516-51. doi: 10.1021/acs.chemrev.5b00562. Epub 2016 Jan 25. Chem Rev. 2016. PMID: 26807783 Free PMC article. Review.
-
Photophysics of fluorescent probes for single-molecule biophysics and super-resolution imaging.Annu Rev Phys Chem. 2012;63:595-617. doi: 10.1146/annurev-physchem-032210-103340. Epub 2012 Jan 30. Annu Rev Phys Chem. 2012. PMID: 22404588 Free PMC article. Review.
-
Single-molecule investigation of substrate binding kinetics and protein conformational dynamics of a B-family replicative DNA polymerase.J Biol Chem. 2013 Apr 19;288(16):11590-600. doi: 10.1074/jbc.M113.459982. Epub 2013 Mar 5. J Biol Chem. 2013. PMID: 23463511 Free PMC article.
-
Efficient pre-catalytic conformational change of reverse transcriptases from SAMHD1 non-counteracting primate lentiviruses during dNTP incorporation.Virology. 2019 Nov;537:36-44. doi: 10.1016/j.virol.2019.08.010. Epub 2019 Aug 14. Virology. 2019. PMID: 31442614 Free PMC article.
-
Structural and Molecular Kinetic Features of Activities of DNA Polymerases.Int J Mol Sci. 2022 Jun 7;23(12):6373. doi: 10.3390/ijms23126373. Int J Mol Sci. 2022. PMID: 35742812 Free PMC article. Review.
References
-
- Joyce CM, Benkovic SJ. DNA polymerase fidelity: kinetics, structure, and checkpoints. Biochemistry. 2004;43:14317–14324. - PubMed
-
- Doublié S, Sawaya MR, Ellenberger T. An open and closed case for all polymerases. Structure. 1999;7:R31–R35. - PubMed
-
- Rothwell PJ, Mitaksov V, Waksman G. Motions of the fingers subdomain of Klentaq1 are fast and not rate limiting: implications for the molecular basis of fidelity in DNA polymerases. Mol Cell. 2005;19:345–355. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
