

Disruption of TAK1 in hepatocytes causes hepatic injury, inflammation, fibrosis, and carcinogenesis
- PMID: 20080763
- PMCID: PMC2818947
- DOI: 10.1073/pnas.0909781107
Disruption of TAK1 in hepatocytes causes hepatic injury, inflammation, fibrosis, and carcinogenesis
Abstract
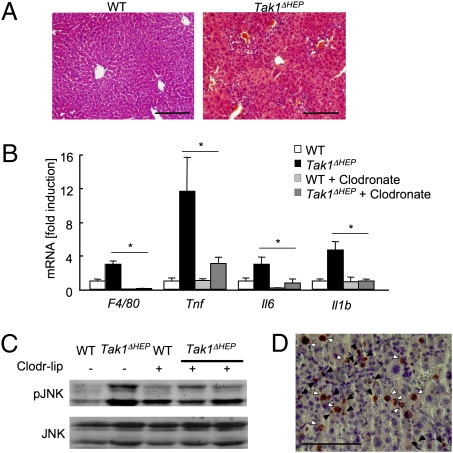
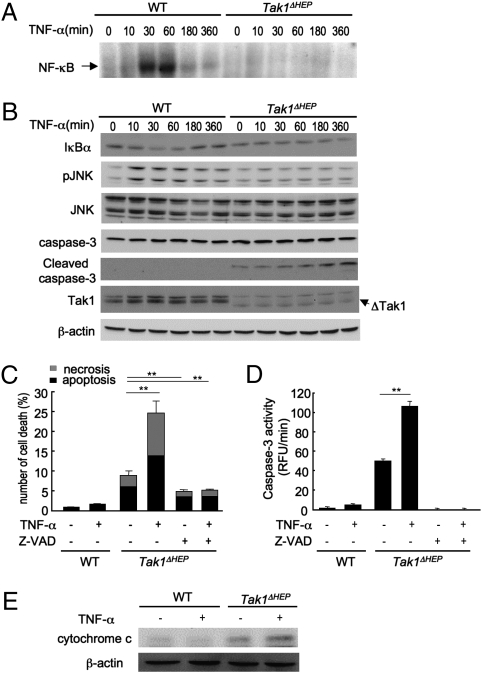
TGF-beta-activated kinase 1 (TAK1) is a MAP3K family member that activates NF-kappaB and JNK via Toll-like receptors and the receptors for IL-1, TNF-alpha, and TGF-beta. Because the TAK1 downstream molecules NF-kappaB and JNK have opposite effects on cell death and carcinogenesis, the role of TAK1 in the liver is unpredictable. To address this issue, we generated hepatocyte-specific Tak1-deficient (Tak1DeltaHEP) mice. The Tak1DeltaHEP mice displayed spontaneous hepatocyte death, compensatory proliferation, inflammatory cell infiltration, and perisinusoidal fibrosis at age 1 month. Older Tak1DeltaHEP mice developed multiple cancer nodules characterized by increased expression of fetal liver genes including alpha-fetoprotein. Cultures of primary hepatocytes deficient in Tak1 exhibited spontaneous cell death that was further increased in response to TNF-alpha. TNF-alpha increased caspase-3 activity but activated neither NF-kappaB nor JNK in Tak1-deficient hepatocytes. Genetic abrogation of TNF receptor type I (TNFRI) in Tak1DeltaHEP mice reduced liver damage, inflammation, and fibrosis compared with unmodified Tak1DeltaHEP mice. In conclusion, hepatocyte-specific deletion of TAK1 in mice resulted in spontaneous hepatocyte death, inflammation, fibrosis, and carcinogenesis that was partially mediated by TNFR signaling, indicating that TAK1 is an essential component for cellular homeostasis in the liver.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Comment in
-
The MAP3K TAK1: a chock block to liver cancer formation.Hepatology. 2010 Oct;52(4):1506-9. doi: 10.1002/hep.23878. Hepatology. 2010. PMID: 20879030 No abstract available.
References
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4:499–511. - PubMed
-
- Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell. 2002;109(Suppl):S81–S96. - PubMed
-
- Maeda S, Kamata H, Luo JL, Leffert H, Karin M. IKKbeta couples hepatocyte death to cytokine-driven compensatory proliferation that promotes chemical hepatocarcinogenesis. Cell. 2005;121:977–990. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
