

Measurement of the membrane curvature preference of phospholipids reveals only weak coupling between lipid shape and leaflet curvature
- PMID: 20080790
- PMCID: PMC2797532
- DOI: 10.1073/pnas.0907354106
Measurement of the membrane curvature preference of phospholipids reveals only weak coupling between lipid shape and leaflet curvature
Abstract
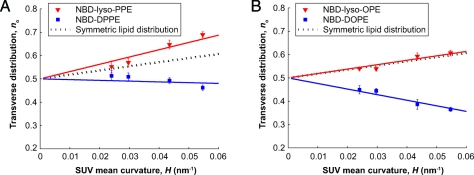
In biological processes, such as fission, fusion and trafficking, it has been shown that lipids of different shapes are sorted into regions with different membrane curvatures. This lipid sorting has been hypothesized to be due to the coupling between the membrane curvature and the lipid's spontaneous curvature, which is related to the lipid's molecular shape. On the other hand, theoretical predictions and simulations suggest that the curvature preference of lipids, due to shape alone, is weaker than that observed in biological processes. To distinguish between these different views, we have directly measured the curvature preferences of several lipids by using a fluorescence-based method. We prepared small unilamellar vesicles of different sizes with a mixture of egg-PC and a small mole fraction of N-nitrobenzoxadiazole (NBD)-labeled phospholipids or lysophospholipids of different chain lengths and saturation, and measured the NBD equilibrium distribution across the bilayer. We observed that the transverse lipid distributions depended linearly on membrane curvature, allowing us to measure the curvature coupling coefficient. Our measurements are in quantitative agreement with predictions based on earlier measurements of the spontaneous curvatures of the corresponding nonfluorescent lipids using X-ray diffraction. We show that, though some lipids have high spontaneous curvatures, they nevertheless showed weak curvature preferences because of the low values of the lipid molecular areas. The weak curvature preference implies that the asymmetric lipid distributions found in biological membranes are not likely to be driven by the spontaneous curvature of the lipids, nor are lipids discriminating sensors of membrane curvature.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Asymmetric distribution of cone-shaped lipids in a highly curved bilayer revealed by a small angle neutron scattering technique.J Phys Condens Matter. 2011 Jul 20;23(28):284104. doi: 10.1088/0953-8984/23/28/284104. Epub 2011 Jun 27. J Phys Condens Matter. 2011. PMID: 21709321
-
Lipid monolayer spontaneous curvatures: A collection of published values.Chem Phys Lipids. 2021 Sep;239:105117. doi: 10.1016/j.chemphyslip.2021.105117. Epub 2021 Jul 12. Chem Phys Lipids. 2021. PMID: 34265278 Review.
-
7-nitrobenz-2-oxa-1,3-diazole-4-yl-labeled phospholipids in lipid membranes: differences in fluorescence behavior.Biophys J. 1996 Jul;71(1):327-35. doi: 10.1016/S0006-3495(96)79228-7. Biophys J. 1996. PMID: 8804615 Free PMC article.
-
Shape changes of giant liposomes induced by an asymmetric transmembrane distribution of phospholipids.Biophys J. 1992 Feb;61(2):347-57. doi: 10.1016/S0006-3495(92)81841-6. Biophys J. 1992. PMID: 1547324 Free PMC article.
-
Chemistry and biology of N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)-labeled lipids: fluorescent probes of biological and model membranes.Chem Phys Lipids. 1990 Mar;53(1):1-15. doi: 10.1016/0009-3084(90)90128-e. Chem Phys Lipids. 1990. PMID: 2191793 Review.
Cited by
-
Nanoscale membrane curvature sorts lipid phases and alters lipid diffusion.Biophys J. 2023 Jun 6;122(11):2203-2215. doi: 10.1016/j.bpj.2023.01.001. Epub 2023 Jan 4. Biophys J. 2023. PMID: 36604961 Free PMC article.
-
Membrane curvature enables N-Ras lipid anchor sorting to liquid-ordered membrane phases.Nat Chem Biol. 2015 Mar;11(3):192-4. doi: 10.1038/nchembio.1733. Epub 2015 Jan 26. Nat Chem Biol. 2015. PMID: 25622090
-
Molecular Shape Solution for Mesoscopic Remodeling of Cellular Membranes.Annu Rev Biophys. 2022 May 9;51:473-497. doi: 10.1146/annurev-biophys-011422-100054. Epub 2022 Mar 3. Annu Rev Biophys. 2022. PMID: 35239417 Free PMC article. Review.
-
Reorganization of Ternary Lipid Mixtures of Nonphosphorylated Phosphatidylinositol Interacting with Angiomotin.J Phys Chem B. 2018 Sep 6;122(35):8404-8415. doi: 10.1021/acs.jpcb.7b12641. Epub 2018 Aug 27. J Phys Chem B. 2018. PMID: 29877706 Free PMC article.
-
Tubular membrane formation of binary giant unilamellar vesicles composed of cylinder and inverse-cone-shaped lipids.Biophys J. 2013 Nov 5;105(9):2074-81. doi: 10.1016/j.bpj.2013.09.021. Biophys J. 2013. PMID: 24209852 Free PMC article.
References
-
- Zimmerberg J, Kozlov MM. How proteins produce cellular membrane curvature. Nat Rev Mol Cell Biol. 2006;7:9–19. - PubMed
-
- Huttner WB, Zimmerberg J. Implications of lipid microdomains for membrane curvature, budding and fission. Curr Opin Cell Biol. 2001;13:478–484. - PubMed
-
- McMahon HT, Gallop JL. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature. 2005;438:590–596. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
