

Overexpression of Osta-siR2141 caused abnormal polarity establishment and retarded growth in rice
- PMID: 20080824
- PMCID: PMC2852654
- DOI: 10.1093/jxb/erp378
Overexpression of Osta-siR2141 caused abnormal polarity establishment and retarded growth in rice
Abstract
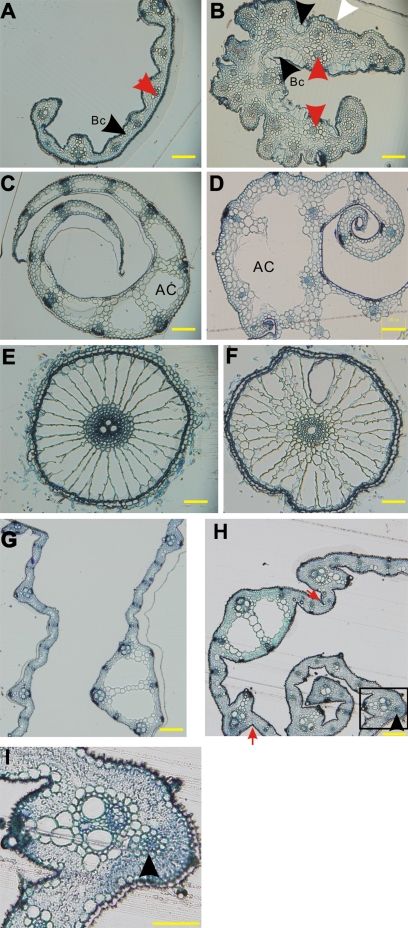
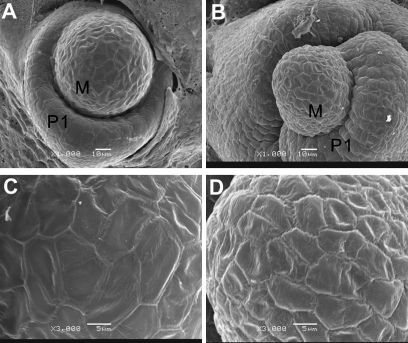
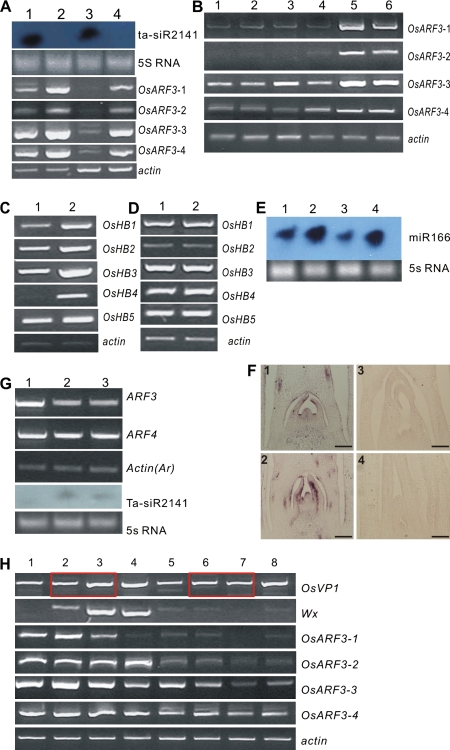
Small RNAs (smRNAs) including miRNAs and siRNAs are critical for gene regulation and plant development. Among the highly diverse siRNAs, trans-acting siRNAs (ta-siRNAs) have been shown to be plant-specific. In Arabidopsis, eight TAS loci belonging to four families (TAS1, TAS2, TAS3, and TAS4) have been identified, and bioinformatics analysis reveals that the sequence of TAS3 is highly conserved in plants. In this study, the function of TAS3 ta-siRNA (tasiR-ARF) has been revealed in rice (Oryza sativa L.) on polarity establishment and stage transition from vegetative to reproductive development by over-expressing Osta-siR2141. Osta-siR2141 replaced miR390 in the miR390 backbone for ectopic expression in rice, and overexpression of Osta-siR2141 caused disturbed vascular bundle development and adaxialization in polarity establishment. Transgenic lines also displayed abnormal shoot apical meristems (SAMs) and retarded growth at the vegetative stage. Molecular analysis revealed that overexpression of Osta-siR2141 resulted in the down-regulation of miR166 and the up-regulation of class III homeodomain-leucine zipper genes (HD-ZIPIIIs) in the vegetative stage but not in the reproductive stage. Moreover, overexpression of Osta-siR2141 in Arabidopsis disturbed polarity establishment and retarded stage transition, suggesting that tasiR-ARF was functionally conserved in rice and Arabidopsis.
Figures




Similar articles
-
TRANS-ACTING SIRNA3-derived short interfering RNAs confer cleavage of mRNAs in rice.Plant Physiol. 2022 Jan 20;188(1):347-362. doi: 10.1093/plphys/kiab452. Plant Physiol. 2022. PMID: 34599593 Free PMC article.
-
Functional diversification of CLAVATA3-related CLE proteins in meristem maintenance in rice.Plant Cell. 2008 Aug;20(8):2049-58. doi: 10.1105/tpc.107.057257. Epub 2008 Aug 1. Plant Cell. 2008. PMID: 18676878 Free PMC article.
-
WAVY LEAF1, an ortholog of Arabidopsis HEN1, regulates shoot development by maintaining MicroRNA and trans-acting small interfering RNA accumulation in rice.Plant Physiol. 2010 Nov;154(3):1335-46. doi: 10.1104/pp.110.160234. Epub 2010 Aug 30. Plant Physiol. 2010. PMID: 20805329 Free PMC article.
-
Transcriptional, post-transcriptional and post-translational regulations of gene expression during leaf polarity formation.Cell Res. 2007 Jun;17(6):512-9. doi: 10.1038/cr.2007.45. Cell Res. 2007. PMID: 17549070 Review.
-
Grass meristems I: shoot apical meristem maintenance, axillary meristem determinacy and the floral transition.Plant Cell Physiol. 2013 Mar;54(3):302-12. doi: 10.1093/pcp/pct025. Epub 2013 Feb 14. Plant Cell Physiol. 2013. PMID: 23411664 Review.
Cited by
-
OsMADS1 Represses microRNA172 in Elongation of Palea/Lemma Development in Rice.Front Plant Sci. 2016 Dec 20;7:1891. doi: 10.3389/fpls.2016.01891. eCollection 2016. Front Plant Sci. 2016. PMID: 28066457 Free PMC article.
-
OsmiR396/growth regulating factor modulate rice grain size through direct regulation of embryo-specific miR408.Plant Physiol. 2021 May 27;186(1):519-533. doi: 10.1093/plphys/kiab084. Plant Physiol. 2021. PMID: 33620493 Free PMC article.
-
A Novel LncRNA, MuLnc1, Associated With Environmental Stress in Mulberry (Morus multicaulis).Front Plant Sci. 2018 May 29;9:669. doi: 10.3389/fpls.2018.00669. eCollection 2018. Front Plant Sci. 2018. PMID: 29896205 Free PMC article.
-
DEGENERATED LEMMA (DEL) regulates lemma development and affects rice grain yield.Physiol Mol Biol Plants. 2023 Mar;29(3):335-347. doi: 10.1007/s12298-023-01297-6. Epub 2023 Mar 23. Physiol Mol Biol Plants. 2023. PMID: 37033767 Free PMC article.
-
OsRAMOSA2 Shapes Panicle Architecture through Regulating Pedicel Length.Front Plant Sci. 2017 Sep 12;8:1538. doi: 10.3389/fpls.2017.01538. eCollection 2017. Front Plant Sci. 2017. PMID: 28955349 Free PMC article.
References
-
- Adenot X, Elmayan T, Lauressergues D, Boutet S, Bouche N, Gasciolli V, Vaucheret H. DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Current Biology. 2006;16:927–932. - PubMed
-
- Allen E, Xie Z, Gustafson AM, Carrington JC. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
-
- Allshire R. Molecular biology. RNAi and heterochromatin: a hushed-up affair. Science. 2002;297:1818–1819. - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, Snyder B, Gaasterland T, Meyer J, Tuschl T. The small RNA profile during Drosophila melanogaster development. Developmental Cell. 2003;5:337–350. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
