

Leaf lifetime photosynthetic rate and leaf demography in whole plants of Ipomoea pes-caprae growing with a low supply of calcium, a 'non-mobile' nutrient
- PMID: 20080828
- PMCID: PMC2814114
- DOI: 10.1093/jxb/erp351
Leaf lifetime photosynthetic rate and leaf demography in whole plants of Ipomoea pes-caprae growing with a low supply of calcium, a 'non-mobile' nutrient
Abstract
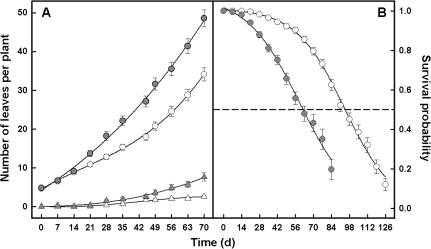
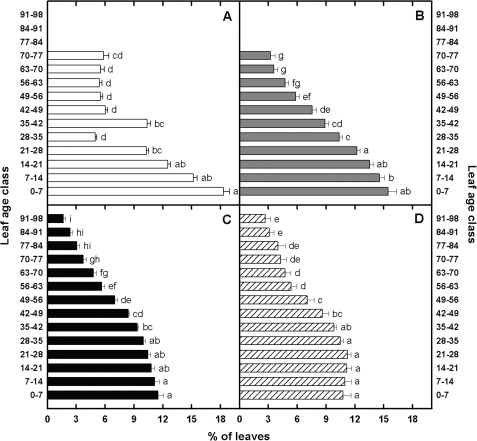
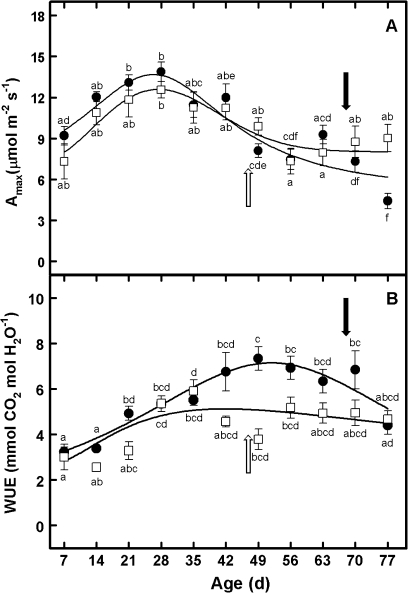
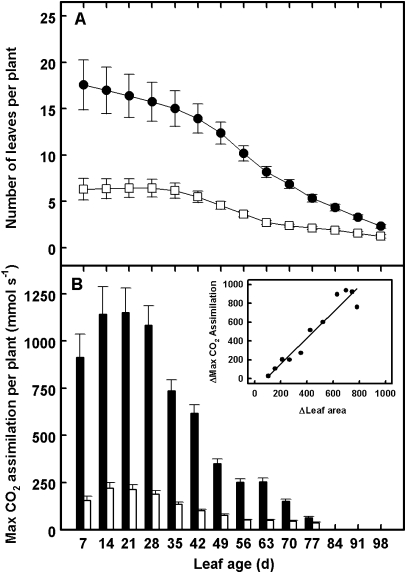
The adaptive significance of leaf longevity has been established in relation to restrictive nutrients that can be retranslocated within the plant. However, the effect of deficiencies in 'non-mobile' nutrients on leaf lifespan and photosynthetic carbon gain is uncertain. Calcium is frequently given as an example of an essential nutrient with low phloem mobility that may alter the leaf senescence process. This study has been designed to estimate leaf lifespan, leaf production (L(p)) and leaf death (L(d)) rates, the age structure of leaves, and the decline in maximum photosynthetic rate (A(max)) with age in plants of Ipomoea pes-caprae growing with a full supply of nutrients and with a low Ca supply. The Ca deficiency produced reductions in L(p) and leaf lifespan compared with control plants. In spite of the differences in the demographic parameters between treatments in control and low-Ca plants, the percentage of leaves of a given leaf age class is maintained in such a way that the number of leaves per plant continues to increase. No relationship was found between Ca supply and A(max). However, the decline in A(max) with leaf senescence was rather sudden in control plants compared with plants growing with a low Ca supply. The importance of simultaneously using the total leaf demographic census and the assimilation rate along with leaf lifespan data in order to understand the performance of whole plants under constrained conditions is discussed.
Figures

Comment in
-
On how to disentangle the contribution of different organs and processes to the growth of whole plants.J Exp Bot. 2010 Mar;61(3):626-8. doi: 10.1093/jxb/erp398. J Exp Bot. 2010. PMID: 20118495 No abstract available.
Similar articles
-
Contributions of leaf photosynthetic capacity, leaf angle and self-shading to the maximization of net photosynthesis in Acer saccharum: a modelling assessment.Ann Bot. 2012 Aug;110(3):731-41. doi: 10.1093/aob/mcs106. Epub 2012 Jun 4. Ann Bot. 2012. PMID: 22665700 Free PMC article.
-
Growth, carbon dioxide exchange and mineral accumulation in potatoes grown at different magnesium concentrations.J Plant Nutr. 1992;15(9):1359-71. doi: 10.1080/01904169209364403. J Plant Nutr. 1992. PMID: 11537503
-
Carbon dioxide enrichment does not reduce leaf longevity or alter accumulation of carbon reserves in the woodland spring ephemeral Erythronium americanum.Ann Bot. 2008 Nov;102(5):835-43. doi: 10.1093/aob/mcn161. Epub 2008 Aug 28. Ann Bot. 2008. PMID: 18757450 Free PMC article.
-
Carbon and nitrogen assimilation in relation to yield: mechanisms are the key to understanding production systems.J Exp Bot. 2002 Apr;53(370):773-87. J Exp Bot. 2002. PMID: 11912221 Review.
-
N uptake and distribution in crops: an agronomical and ecophysiological perspective.J Exp Bot. 2002 Apr;53(370):789-99. doi: 10.1093/jexbot/53.370.789. J Exp Bot. 2002. PMID: 11912222 Review.
References
-
- Ackerly DD. Self-shading, carbon gain and leaf dynamics: a test of alternative optimality models. Oecologia. 1999;119:300–310. - PubMed
-
- Aerts R. Nutrient resorption from senescing leaves of perennials: are there general patterns? Journal of Ecology. 1996;84:597–608.
-
- Aphalo PJ, Schoettle AW, Lehto T. Leaf life span and the mobility of ’non-mobile’" mineral nutrients: the case of boron in conifers. Silva Fennica. 2002;36:671–680.
-
- Bach CE. Effects of clonal integration on response to sand burial and defoliation by the dune plant Ipomoea pes-caprae (Convolvulaceae) Australian Journal of Botany. 2000;48:159–166.
-
- Bazzaz FA, Harper JL. Demographic analysis of the growth of Linum usitatissimum. New Phytologist. 1977;78:193–208.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
