

Silent information regulator 3: the Goldilocks of the silencing complex
- PMID: 20080949
- PMCID: PMC2807346
- DOI: 10.1101/gad.1865510
Silent information regulator 3: the Goldilocks of the silencing complex
Abstract
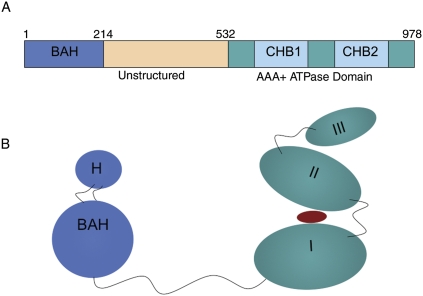
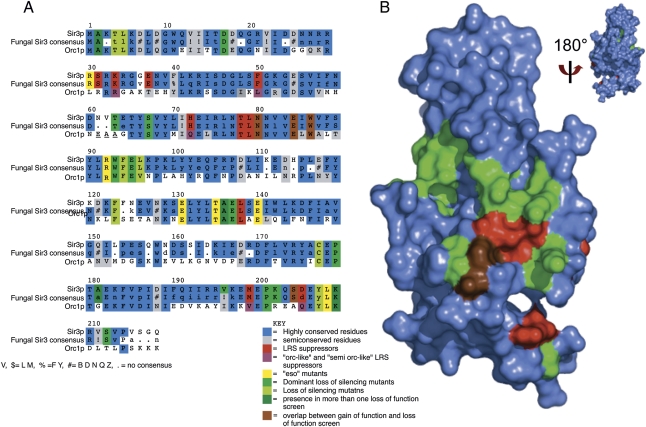
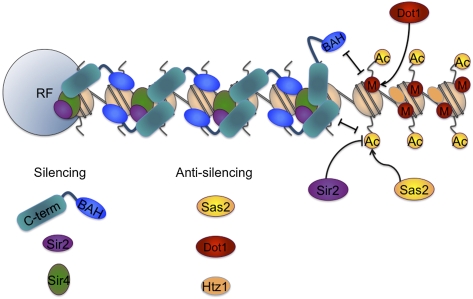
A recent explosion of work surrounds the interactions between Sir3p (Silent Information Regulator 3) and chromatin. We review here the Sir3p functions related to its role in silencing in Saccharomyces cerevisiae. This unusual protein, which is absolutely required for silencing, is distantly related to the highly conserved replication initiator Orc1p, but is itself phylogenetically limited to "post-genome-duplicated" budding yeasts. Several recent studies revise earlier models for Sir3p action. Specifically, the N-terminal bromo-adjacent homology (BAH) domain plays a now well-defined role in silencing, and a picture is emerging in which both termini of Sir3p bind two locations on the nucleosome: (1) the loss of ribosomal DNA silencing (LRS) surface in the nucleosome core, and (2) the N-terminal histone tails for effective silencing at telomeres. We relate Sir3p structure and function, and summarize recent molecular studies of Sir3p/chromatin binding, Sir3p/Dot1p competition, and the possible role of O-Acetyl ADP ribose (O-AADPR) in Sir3p/chromatin binding. We emphasize recent genetic data that provide important new insights and settle controversies created by in vitro work. Finally, we synthesize these ideas to revise the model for how Sir3p mediates silent chromatin formation in yeast, in part through its affinity for the LRS region of the nucleosome, which must be "just right."
Figures
Similar articles
-
Compensatory interactions between Sir3p and the nucleosomal LRS surface imply their direct interaction.PLoS Genet. 2008 Dec;4(12):e1000301. doi: 10.1371/journal.pgen.1000301. Epub 2008 Dec 12. PLoS Genet. 2008. PMID: 19079580 Free PMC article.
-
A region of the nucleosome required for multiple types of transcriptional silencing in Saccharomyces cerevisiae.Genetics. 2011 Jul;188(3):535-48. doi: 10.1534/genetics.111.129197. Epub 2011 May 5. Genetics. 2011. PMID: 21546544 Free PMC article.
-
Structure and function of the BAH-containing domain of Orc1p in epigenetic silencing.EMBO J. 2002 Sep 2;21(17):4600-11. doi: 10.1093/emboj/cdf468. EMBO J. 2002. PMID: 12198162 Free PMC article.
-
Silent chromatin in yeast: an orchestrated medley featuring Sir3p [corrected].Bioessays. 1998 Jan;20(1):30-40. doi: 10.1002/(SICI)1521-1878(199801)20:1<30::AID-BIES6>3.0.CO;2-W. Bioessays. 1998. PMID: 9504045 Review.
-
Function and metabolism of sirtuin metabolite O-acetyl-ADP-ribose.Biochim Biophys Acta. 2010 Aug;1804(8):1617-25. doi: 10.1016/j.bbapap.2010.02.007. Epub 2010 Feb 20. Biochim Biophys Acta. 2010. PMID: 20176146 Free PMC article. Review.
Cited by
-
Detection of an altered heterochromatin structure in the absence of the nucleotide excision repair protein Rad4 in Saccharomyces cerevisiae.Cell Cycle. 2013 Aug 1;12(15):2435-42. doi: 10.4161/cc.25457. Epub 2013 Jul 3. Cell Cycle. 2013. PMID: 23839037 Free PMC article.
-
Structure and function of the Orc1 BAH-nucleosome complex.Nat Commun. 2019 Jul 1;10(1):2894. doi: 10.1038/s41467-019-10609-y. Nat Commun. 2019. PMID: 31263106 Free PMC article.
-
Dynamics of Sir3 spreading in budding yeast: secondary recruitment sites and euchromatic localization.EMBO J. 2011 Mar 16;30(6):1012-26. doi: 10.1038/emboj.2011.30. Epub 2011 Feb 18. EMBO J. 2011. PMID: 21336256 Free PMC article.
-
Epigenetic silencing mediates mitochondria stress-induced longevity.Cell Metab. 2013 Jun 4;17(6):954-964. doi: 10.1016/j.cmet.2013.04.003. Cell Metab. 2013. PMID: 23747251 Free PMC article.
-
Selection of Protein-Protein Interactions of Desired Affinities with a Bandpass Circuit.J Mol Biol. 2019 Jan 18;431(2):391-400. doi: 10.1016/j.jmb.2018.11.011. Epub 2018 Nov 15. J Mol Biol. 2019. PMID: 30448232 Free PMC article.
References
-
- Callebaut I, Courvalin JC, Mornon JP. The BAH (bromo-adjacent homology) domain: A link between DNA methylation, replication and transcriptional regulation. FEBS Lett. 1999;446:189–193. - PubMed
-
- Chang JF, Hall BE, Tanny JC, Moazed D, Filman D, Ellenberger T. Structure of the coiled-coil dimerization motif of Sir4 and its interaction with Sir3. Structure. 2003;11:637–649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases