

PcaO positively regulates pcaHG of the beta-ketoadipate pathway in Corynebacterium glutamicum
- PMID: 20081038
- PMCID: PMC2832523
- DOI: 10.1128/JB.01338-09
PcaO positively regulates pcaHG of the beta-ketoadipate pathway in Corynebacterium glutamicum
Abstract
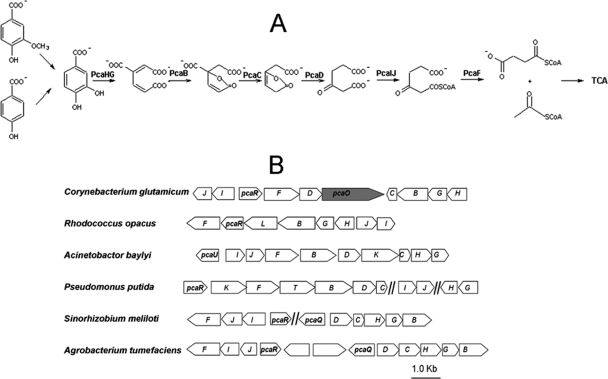
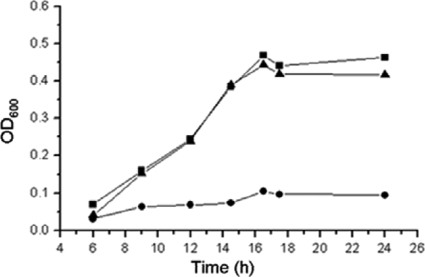
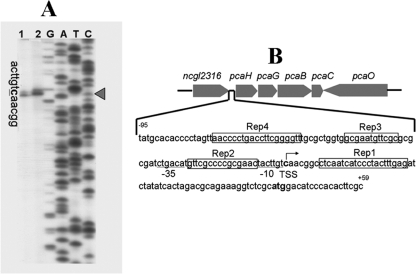
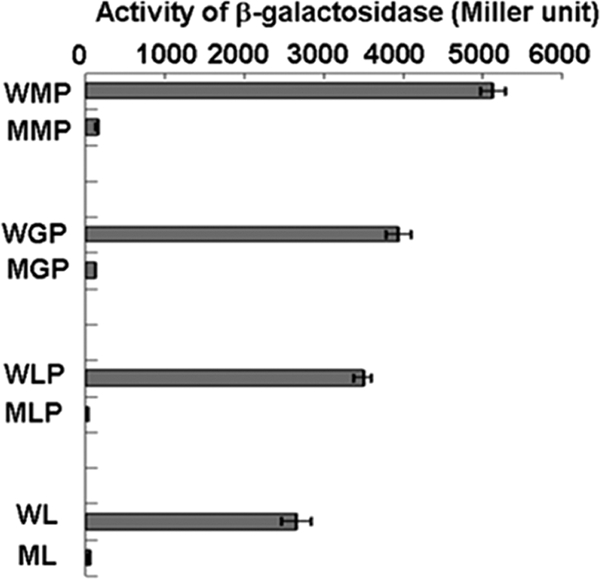
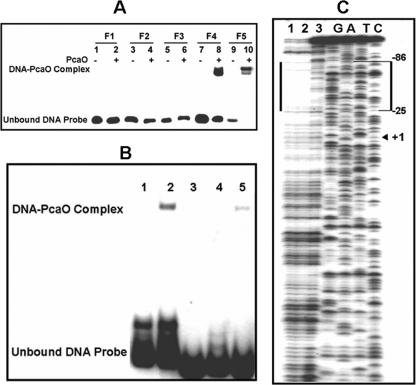
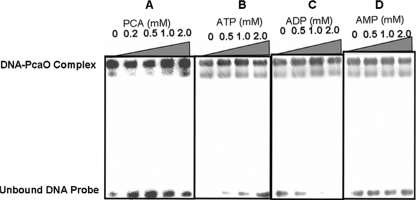
We identified a new regulator, PcaO, which is involved in regulation of the protocatechuate (PCA) branch of the beta-ketoadipate pathway in Corynebacterium glutamicum. PcaO is an atypical large ATP-binding LuxR family (LAL)-type regulator and does not have a Walker A motif. A mutant of C. glutamicum in which pcaO was disrupted (RES167DeltapcaO) was unable to grow on PCA, and growth on PCA was restored by complementation with pcaO. Both an enzymatic assay of PCA 3,4-dioxygenase activity (encoded by pcaHG) and transcriptional analysis of pcaHG by reverse transcription-PCR revealed that PcaO positively regulated pcaHG. A promoter-LacZ transcriptional fusion assay suggested that PcaO interacted with the sequence upstream of pcaHG. Electrophoretic mobility shift assay (EMSA) analysis indicated that an imperfect palindromic sequence ((-78)AACCCCTGACCTTCGGGGTT(-59)) that was located upstream of the -35 region of the pcaHG promoter was essential for PcaO regulation. DNase I footprinting showed that this imperfect palindrome was protected from DNase I digestion. Site-directed mutation and EMSA tests revealed that this palindrome sequence was essential for PcaO binding to the DNA fragment. In vitro EMSA results showed that ATP weakened the binding between PcaO and its target sequence but ADP strengthened this binding, while the effect of protocatechuate on PcaO binding was dependent on the protocatechuate concentration.
Figures
Similar articles
-
Key enzymes of the protocatechuate branch of the beta-ketoadipate pathway for aromatic degradation in Corynebacterium glutamicum.Sci China C Life Sci. 2005 Jun;48(3):241-9. doi: 10.1007/BF03183617. Sci China C Life Sci. 2005. PMID: 16092756
-
Involvement of the global regulator GlxR in 3-hydroxybenzoate and gentisate utilization by Corynebacterium glutamicum.Appl Environ Microbiol. 2014 Jul;80(14):4215-25. doi: 10.1128/AEM.00290-14. Epub 2014 May 2. Appl Environ Microbiol. 2014. PMID: 24795375 Free PMC article.
-
GenR, an IclR-type regulator, activates and represses the transcription of gen genes involved in 3-hydroxybenzoate and gentisate catabolism in Corynebacterium glutamicum.J Bacteriol. 2013 Apr;195(7):1598-609. doi: 10.1128/JB.02216-12. Epub 2013 Jan 25. J Bacteriol. 2013. PMID: 23354754 Free PMC article.
-
Involvement of the LuxR-type transcriptional regulator RamA in regulation of expression of the gapA gene, encoding glyceraldehyde-3-phosphate dehydrogenase of Corynebacterium glutamicum.J Bacteriol. 2009 Feb;191(3):968-77. doi: 10.1128/JB.01425-08. Epub 2008 Dec 1. J Bacteriol. 2009. PMID: 19047347 Free PMC article.
-
Transcriptional regulation of catabolic pathways for aromatic compounds in Corynebacterium glutamicum.Genet Mol Res. 2006 Dec 7;5(4):773-89. Genet Mol Res. 2006. PMID: 17183485 Review.
Cited by
-
Comprehensive discovery and characterization of small RNAs in Corynebacterium glutamicum ATCC 13032.BMC Genomics. 2013 Oct 19;14(1):714. doi: 10.1186/1471-2164-14-714. BMC Genomics. 2013. PMID: 24138339 Free PMC article.
-
Phenylacetic acid catabolism and its transcriptional regulation in Corynebacterium glutamicum.Appl Environ Microbiol. 2012 Aug;78(16):5796-804. doi: 10.1128/AEM.01588-12. Epub 2012 Jun 8. Appl Environ Microbiol. 2012. PMID: 22685150 Free PMC article.
-
Artificial oxidative stress-tolerant Corynebacterium glutamicum.AMB Express. 2014 Mar 18;4:15. doi: 10.1186/s13568-014-0015-1. eCollection 2014. AMB Express. 2014. PMID: 24949252 Free PMC article.
-
Improving growth properties of Corynebacterium glutamicum by implementing an iron-responsive protocatechuate biosynthesis.Microb Biotechnol. 2023 May;16(5):1041-1053. doi: 10.1111/1751-7915.14244. Epub 2023 Mar 11. Microb Biotechnol. 2023. PMID: 36905370 Free PMC article.
-
Microbial single-cell growth response at defined carbon limiting conditions.RSC Adv. 2019 May 7;9(25):14040-14050. doi: 10.1039/c9ra02454a. eCollection 2019 May 7. RSC Adv. 2019. PMID: 35519298 Free PMC article.
References
-
- Brinkrolf, K., I. Brune, and A. Tauch. 2006. Transcriptional regulation of catabolic pathways for aromatic compounds in Corynebacterium glutamicum. Genet. Mol. Res. 5:773-789. - PubMed
-
- Brown, C. K., M. W. Vetting, C. A. Earhart, and D. H. Ohlendorf. 2004. Biophysical analyses of designed and selected mutants of PCA 3,4-dioxygenase. Annu. Rev. Microbiol. 58:555-585. - PubMed
-
- Brzostowicz, P. C., A. B. Reams, T. J. Clark, and E. L. Neidle. 2003. Transcriptional cross-regulation of the catechol and PCA branches of the β-ketoadipate pathway contributes to carbon source-dependent expression of the Acinetobacter sp. strain ADP1 pobA gene. J. Bacteriol. 69:1598-1606. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
