

Bifurcation and enhancement of autonomous-nonautonomous retrotransposon partnership through LTR Swapping in soybean
- PMID: 20081112
- PMCID: PMC2828711
- DOI: 10.1105/tpc.109.068775
Bifurcation and enhancement of autonomous-nonautonomous retrotransposon partnership through LTR Swapping in soybean
Abstract
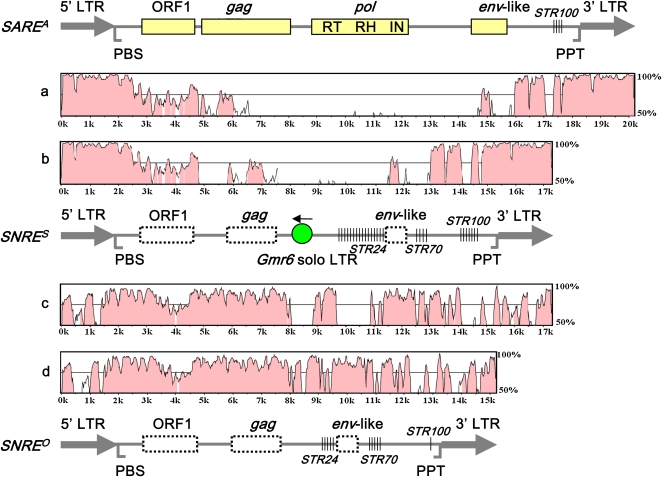
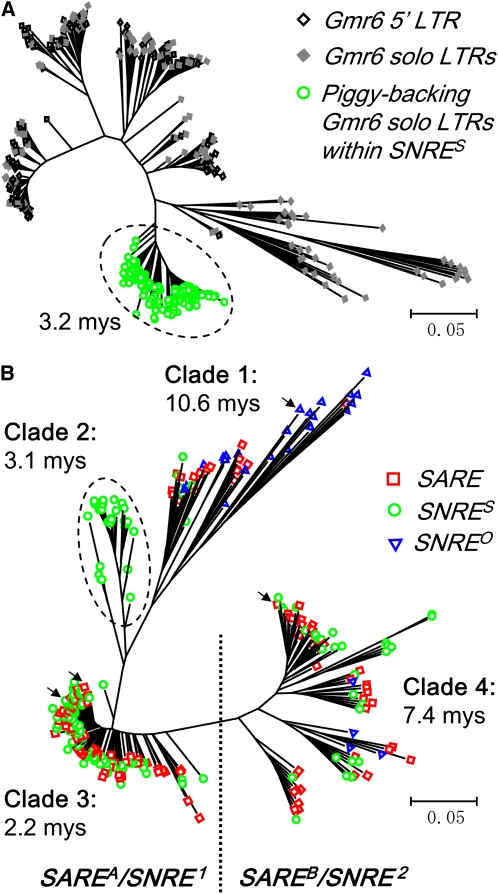
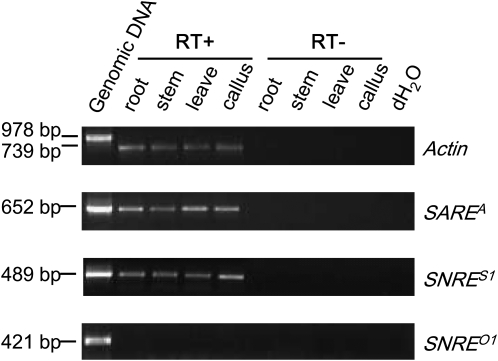
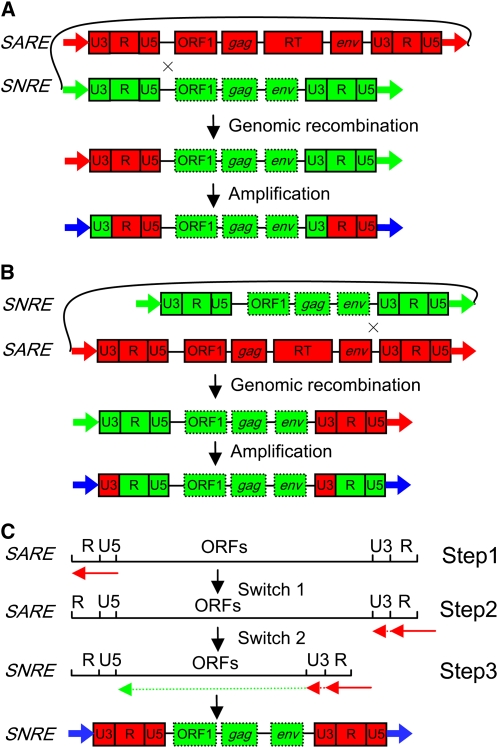
Long terminal repeat (LTR) retrotransposons, the most abundant genomic components in flowering plants, are classifiable into autonomous and nonautonomous elements based on their structural completeness and transposition capacity. It has been proposed that selection is the major force for maintaining sequence (e.g., LTR) conservation between nonautonomous elements and their autonomous counterparts. Here, we report the structural, evolutionary, and expression characterization of a giant retrovirus-like soybean (Glycine max) LTR retrotransposon family, SNARE. This family contains two autonomous subfamilies, SARE(A) and SARE(B), that appear to have evolved independently since the soybean genome tetraploidization event approximately 13 million years ago, and a nonautonomous subfamily, SNRE, that originated from SARE(A). Unexpectedly, a subset of the SNRE elements, which amplified from a single founding SNRE element within the last approximately 3 million years, have been dramatically homogenized with either SARE(A) or SARE(B) primarily in the LTR regions and bifurcated into distinct subgroups corresponding to the two autonomous subfamilies. We uncovered evidence of region-specific swapping of nonautonomous elements with autonomous elements that primarily generated various nonautonomous recombinants with LTR sequences from autonomous elements of different evolutionary lineages, thus revealing a molecular mechanism for the enhancement of preexisting partnership and the establishment of new partnership between autonomous and nonautonomous elements.
Figures



Similar articles
-
Replication of nonautonomous retroelements in soybean appears to be both recent and common.Plant Physiol. 2008 Dec;148(4):1760-71. doi: 10.1104/pp.108.127910. Epub 2008 Oct 24. Plant Physiol. 2008. PMID: 18952860 Free PMC article.
-
Evolutionary conservation, diversity and specificity of LTR-retrotransposons in flowering plants: insights from genome-wide analysis and multi-specific comparison.Plant J. 2010 Aug;63(4):584-98. doi: 10.1111/j.1365-313X.2010.04263.x. Plant J. 2010. PMID: 20525006
-
Analyses of LTR-retrotransposon structures reveal recent and rapid genomic DNA loss in rice.Genome Res. 2004 May;14(5):860-9. doi: 10.1101/gr.1466204. Epub 2004 Apr 12. Genome Res. 2004. PMID: 15078861 Free PMC article.
-
[Non-LTR retrotransposons: LINEs and SINEs in plant genome].Yi Chuan. 2006 Jun;28(6):731-6. Yi Chuan. 2006. PMID: 16818439 Review. Chinese.
-
LTR retrotransposons and flowering plant genome size: emergence of the increase/decrease model.Cytogenet Genome Res. 2005;110(1-4):91-107. doi: 10.1159/000084941. Cytogenet Genome Res. 2005. PMID: 16093661 Review.
Cited by
-
PlantLTRdb: An interactive database for 195 plant species LTR-retrotransposons.Front Plant Sci. 2023 Mar 6;14:1134627. doi: 10.3389/fpls.2023.1134627. eCollection 2023. Front Plant Sci. 2023. PMID: 36950350 Free PMC article.
-
LTR_retriever: A Highly Accurate and Sensitive Program for Identification of Long Terminal Repeat Retrotransposons.Plant Physiol. 2018 Feb;176(2):1410-1422. doi: 10.1104/pp.17.01310. Epub 2017 Dec 12. Plant Physiol. 2018. PMID: 29233850 Free PMC article.
-
High-frequency recombination between members of an LTR retrotransposon family during transposition bursts.Nat Commun. 2017 Nov 3;8(1):1283. doi: 10.1038/s41467-017-01374-x. Nat Commun. 2017. PMID: 29097664 Free PMC article.
-
Retrotransposon domain swapping.Plant Cell. 2010 Jan;22(1):2. doi: 10.1105/tpc.109.220111. Epub 2010 Jan 15. Plant Cell. 2010. PMID: 20081113 Free PMC article. No abstract available.
-
Comparative analysis of miniature inverted-repeat transposable elements (MITEs) and long terminal repeat (LTR) retrotransposons in six Citrus species.BMC Plant Biol. 2019 Apr 15;19(1):140. doi: 10.1186/s12870-019-1757-3. BMC Plant Biol. 2019. PMID: 30987586 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
