

Jasmonate and ppHsystemin regulate key Malonylation steps in the biosynthesis of 17-Hydroxygeranyllinalool Diterpene Glycosides, an abundant and effective direct defense against herbivores in Nicotiana attenuata
- PMID: 20081114
- PMCID: PMC2828710
- DOI: 10.1105/tpc.109.071449
Jasmonate and ppHsystemin regulate key Malonylation steps in the biosynthesis of 17-Hydroxygeranyllinalool Diterpene Glycosides, an abundant and effective direct defense against herbivores in Nicotiana attenuata
Abstract
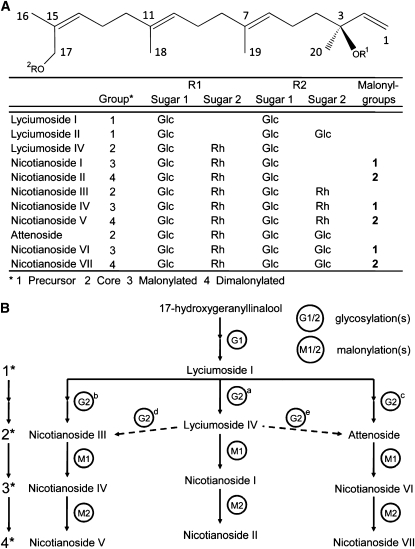
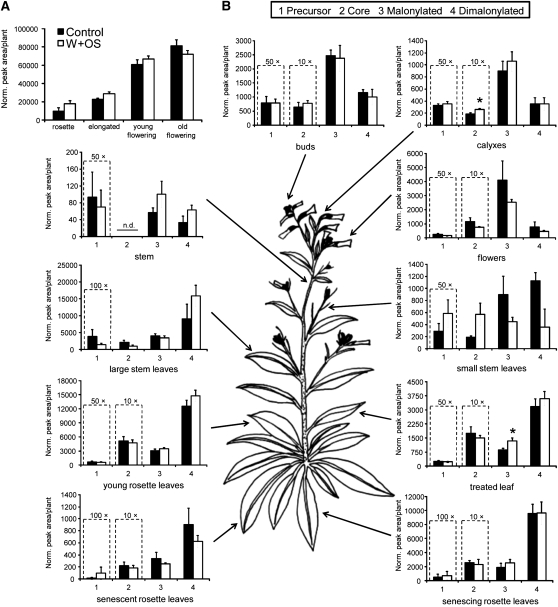
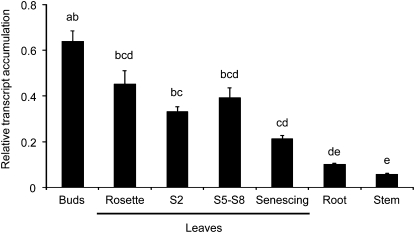
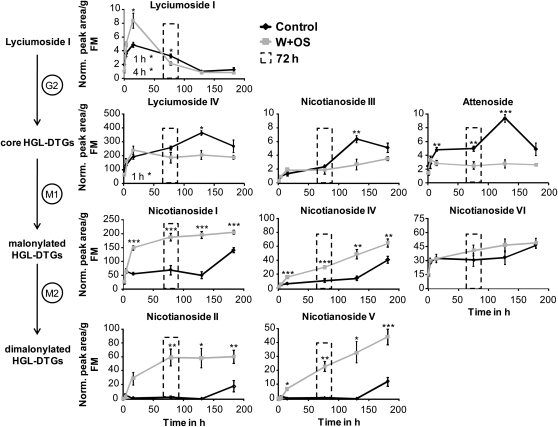
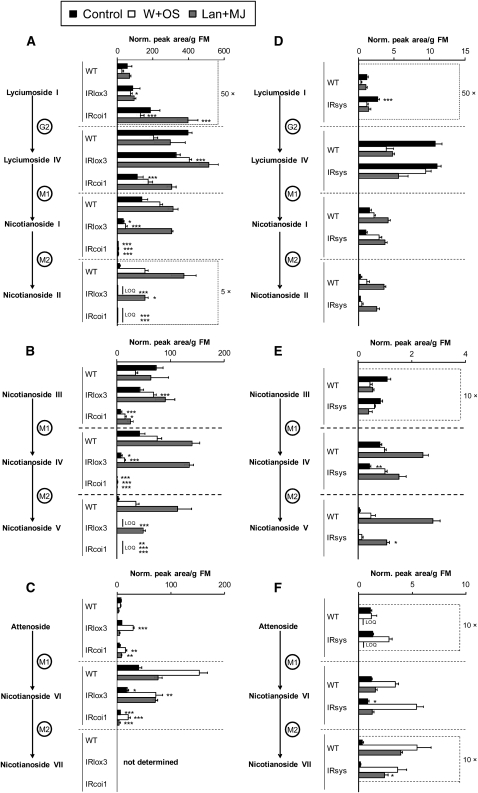
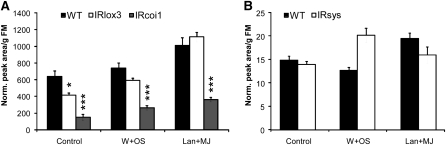
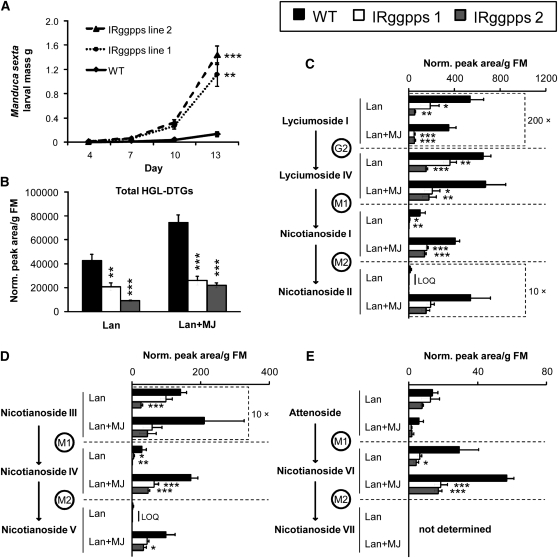
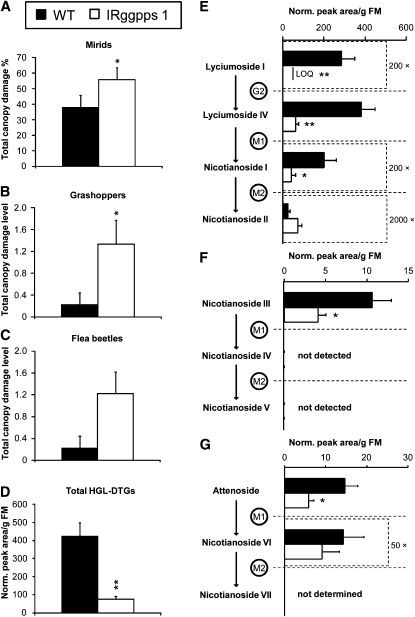
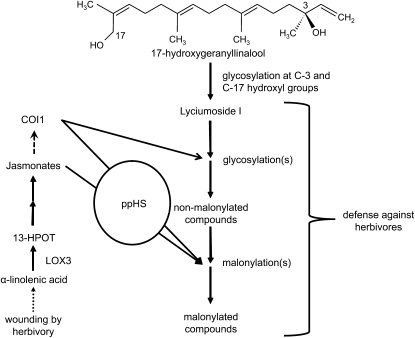
We identified 11 17-hydroxygeranyllinalool diterpene glycosides (HGL-DTGs) that occur in concentrations equivalent to starch (mg/g fresh mass) in aboveground tissues of coyote tobacco (Nicotiana attenuata) and differ in their sugar moieties and malonyl sugar esters (0-2). Concentrations of HGL-DTGs, particularly malonylated compounds, are highest in young and reproductive tissues. Within a tissue, herbivore elicitation changes concentrations and biosynthetic kinetics of individual compounds. Using stably transformed N. attenuata plants silenced in jasmonate production and perception, or production of N. attenuata Hyp-rich glycopeptide systemin precursor by RNA interference, we identified malonylation as the key biosynthetic step regulated by herbivory and jasmonate signaling. We stably silenced N. attenuata geranylgeranyl diphosphate synthase (ggpps) to reduce precursors for the HGL-DTG skeleton, resulting in reduced total HGL-DTGs and greater vulnerability to native herbivores in the field. Larvae of the specialist tobacco hornworm (Manduca sexta) grew up to 10 times as large on ggpps silenced plants, and silenced plants suffered significantly more damage from herbivores in N. attenuata's native habitat than did wild-type plants. We propose that high concentrations of HGL-DTGs effectively defend valuable tissues against herbivores and that malonylation may play an important role in regulating the distribution and storage of HGL-DTGs in plants.
Figures
Similar articles
-
Specific decorations of 17-hydroxygeranyllinalool diterpene glycosides solve the autotoxicity problem of chemical defense in Nicotiana attenuata.Plant Cell. 2021 Jul 2;33(5):1748-1770. doi: 10.1093/plcell/koab048. Plant Cell. 2021. PMID: 33561278 Free PMC article.
-
Silencing geranylgeranyl diphosphate synthase in Nicotiana attenuata dramatically impairs resistance to tobacco hornworm.Plant Physiol. 2008 Mar;146(3):974-86. doi: 10.1104/pp.107.108811. Epub 2007 Oct 26. Plant Physiol. 2008. PMID: 17965175 Free PMC article.
-
UVB radiation and 17-hydroxygeranyllinalool diterpene glycosides provide durable resistance against mirid (Tupiocoris notatus) attack in field-grown Nicotiana attenuata plants.Plant Cell Environ. 2013 Mar;36(3):590-606. doi: 10.1111/j.1365-3040.2012.02598.x. Epub 2012 Sep 18. Plant Cell Environ. 2013. PMID: 22897424
-
The essential role of jasmonic acid in plant-herbivore interactions--using the wild tobacco Nicotiana attenuata as a model.J Genet Genomics. 2013 Dec 20;40(12):597-606. doi: 10.1016/j.jgg.2013.10.001. Epub 2013 Nov 9. J Genet Genomics. 2013. PMID: 24377866 Review.
-
Harmonizing biosynthesis with post-ingestive modifications to understand the ecological functions of plant natural products.Nat Prod Rep. 2022 Jul 20;39(7):1383-1392. doi: 10.1039/d2np00019a. Nat Prod Rep. 2022. PMID: 35575224 Free PMC article. Review.
Cited by
-
Empoasca leafhoppers attack wild tobacco plants in a jasmonate-dependent manner and identify jasmonate mutants in natural populations.Proc Natl Acad Sci U S A. 2012 Jun 12;109(24):E1548-57. doi: 10.1073/pnas.1200363109. Epub 2012 May 21. Proc Natl Acad Sci U S A. 2012. PMID: 22615404 Free PMC article.
-
Strigolactone signaling regulates specialized metabolism in tobacco stems and interactions with stem-feeding herbivores.PLoS Biol. 2020 Aug 18;18(8):e3000830. doi: 10.1371/journal.pbio.3000830. eCollection 2020 Aug. PLoS Biol. 2020. PMID: 32810128 Free PMC article.
-
ZEITLUPE in the Roots of Wild Tobacco Regulates Jasmonate-Mediated Nicotine Biosynthesis and Resistance to a Generalist Herbivore.Plant Physiol. 2018 Jun;177(2):833-846. doi: 10.1104/pp.18.00315. Epub 2018 May 2. Plant Physiol. 2018. PMID: 29720557 Free PMC article.
-
S-Nitrosoglutathione reductase (GSNOR) mediates the biosynthesis of jasmonic acid and ethylene induced by feeding of the insect herbivore Manduca sexta and is important for jasmonate-elicited responses in Nicotiana attenuata.J Exp Bot. 2011 Aug;62(13):4605-16. doi: 10.1093/jxb/err171. Epub 2011 May 27. J Exp Bot. 2011. PMID: 21622839 Free PMC article.
-
The decoration of specialized metabolites influences stylar development.Elife. 2018 Oct 5;7:e38611. doi: 10.7554/eLife.38611. Elife. 2018. PMID: 30289384 Free PMC article.
References
-
- Baldwin I.T. (1996). Allometric limits to the induced accumulation of nicotine in wild tobacco. Plant Species Biol. 11: 107–114
-
- Baldwin I.T. (1999). Inducible nicotine production in native Nicotiana as an example of adaptive phenotypic plasticity. J. Chem. Ecol. 25: 3–30
-
- Baldwin I.T., Schmelz E.A. (1996). Immunological “memory” in the induced accumulation of nicotine in wild tobacco. Ecology 77: 236–246
-
- Baldwin I.T., Zhang Z.-P., Diab N., Ohnmeiss T.E., McCloud E.S., Lynds G.Y., Schmelz E.A. (1997). Quantification, correlations and manipulations of wound-induced changes in jasmonic acid and nicotine in Nicotiana sylvestris.. Planta 201: 397–404
-
- Berger B., Baldwin I.T. (2007). The hydroxyproline-rich glycopeptide systemin precursor NapreproHypSys does not play a central role in Nicotiana attenuata's anti-herbivore defense responses. Plant Cell Environ. 30: 1450–1464 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
