

The EVERSHED receptor-like kinase modulates floral organ shedding in Arabidopsis
- PMID: 20081191
- PMCID: PMC2858908
- DOI: 10.1242/dev.041335
The EVERSHED receptor-like kinase modulates floral organ shedding in Arabidopsis
Abstract
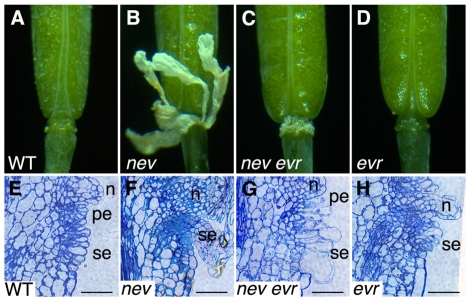
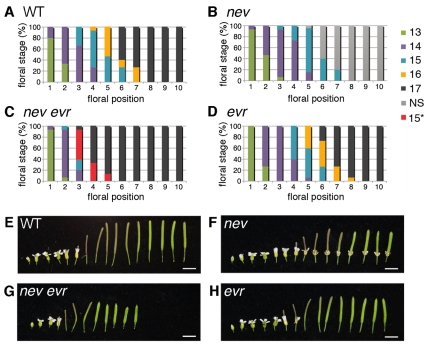
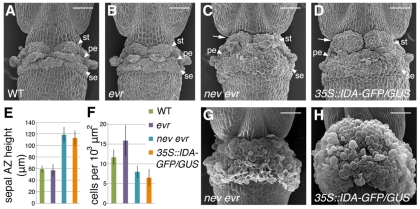
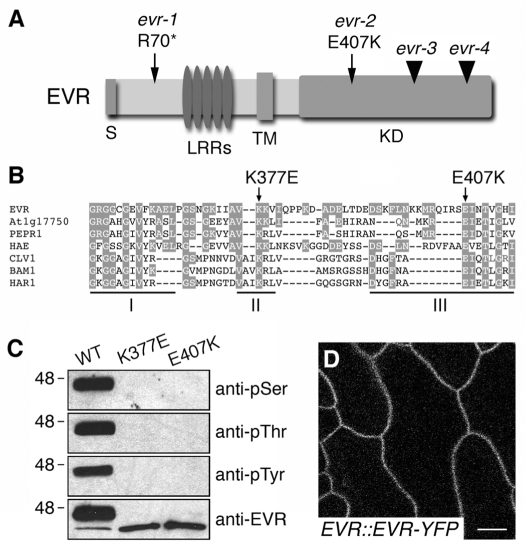
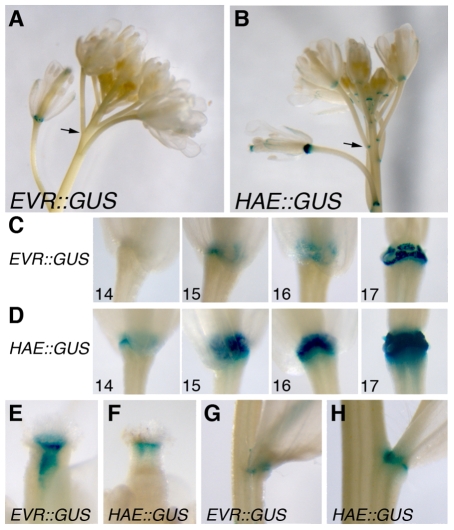
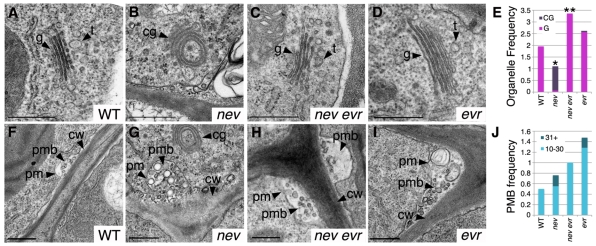
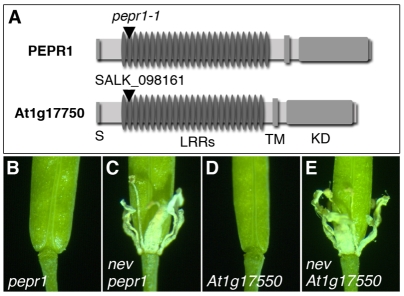
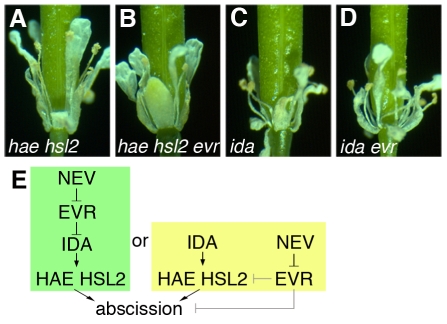
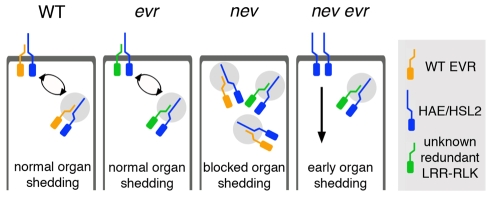
Plant cell signaling triggers the abscission of entire organs, such as fruit, leaves and flowers. Previously, we characterized an ADP-ribosylation factor GTPase-activating protein, NEVERSHED (NEV), that regulates membrane trafficking and is essential for floral organ shedding in Arabidopsis. Through a screen for mutations that restore organ separation in nev flowers, we have identified a leucine-rich repeat receptor-like kinase, EVERSHED (EVR), that functions as an inhibitor of abscission. Defects in the Golgi structure and location of the trans-Golgi network in nev abscission zone cells are rescued by a mutation in EVR, suggesting that EVR might regulate membrane trafficking during abscission. In addition to shedding their floral organs prematurely, nev evr flowers show enlarged abscission zones. A similar phenotype was reported for plants ectopically expressing INFLORESCENCE DEFICIENT IN ABSCISSION, a predicted signaling ligand for the HAESA/HAESA-LIKE2 receptor-like kinases, indicating that this signaling pathway may be constitutively active in nev evr flowers. We present a model in which EVR modulates the timing and region of abscission by promoting the internalization of other receptor-like kinases from the plasma membrane.
Figures
References
-
- Alexandersson E., Saalbach, G., Larsson, C. and Kjellbom, P. (2004). Arabidopsis plasma membrane proteomics identifies components of transport, signal transduction and membrane trafficking. Plant Cell Physiol. 45, 1543-1556. - PubMed
-
- Alonso J. M., Stepanova, A. N., Leisse, T. J., Kim, C. J., Chen, H., Shinn, P., Stevenson, D. K., Zimmerman, J., Barajas, P., Cheuk, R., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657. - PubMed
-
- Ascencio-Ibáñez J. T., Sozzani, R., Lee, T.-J., Chu, T.-M., Wolfinger, R. D., Cella, R. and Hanley-Bowdoin, L. (2008). Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 148, 436-454. - PMC - PubMed
-
- Blázquez M. A., Soowal, L. N., Lee, I. and Weigel, D. (1997). LEAFY expression and flower initiation in Arabidopsis. Development 124, 3835-3844. - PubMed
-
- Boller T. and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60, 379-406. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
