

Early evolution of the LIM homeobox gene family
- PMID: 20082688
- PMCID: PMC2828406
- DOI: 10.1186/1741-7007-8-4
Early evolution of the LIM homeobox gene family
Abstract
Background: LIM homeobox (Lhx) transcription factors are unique to the animal lineage and have patterning roles during embryonic development in flies, nematodes and vertebrates, with a conserved role in specifying neuronal identity. Though genes of this family have been reported in a sponge and a cnidarian, the expression patterns and functions of the Lhx family during development in non-bilaterian phyla are not known.
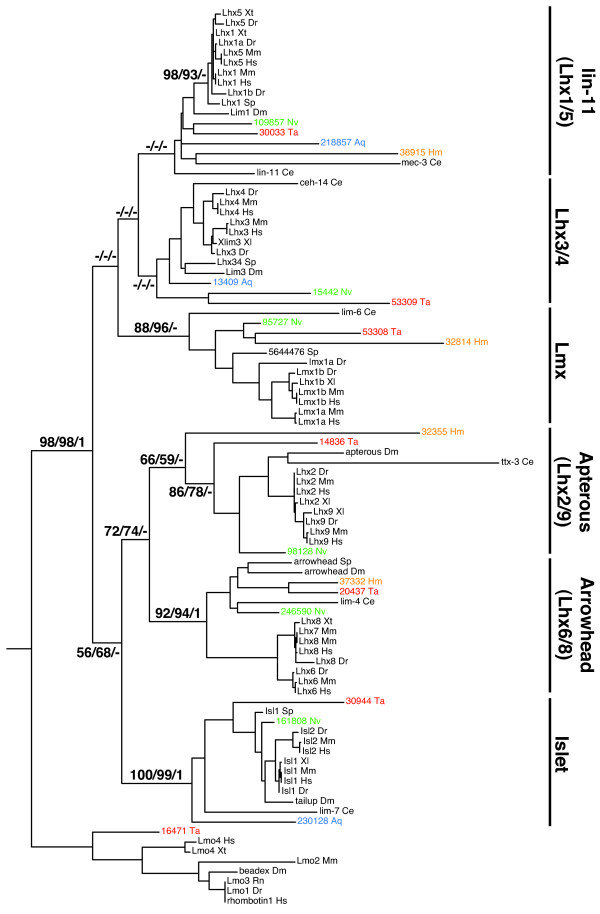
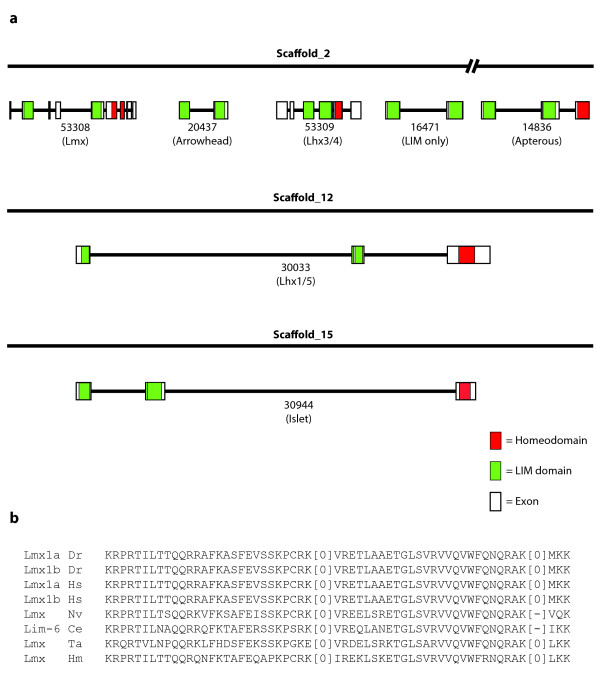
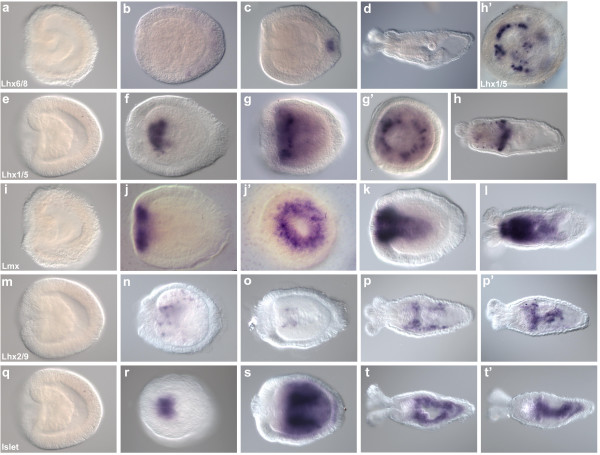
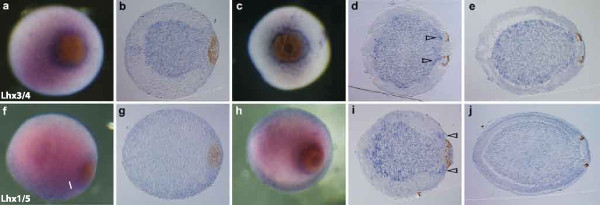
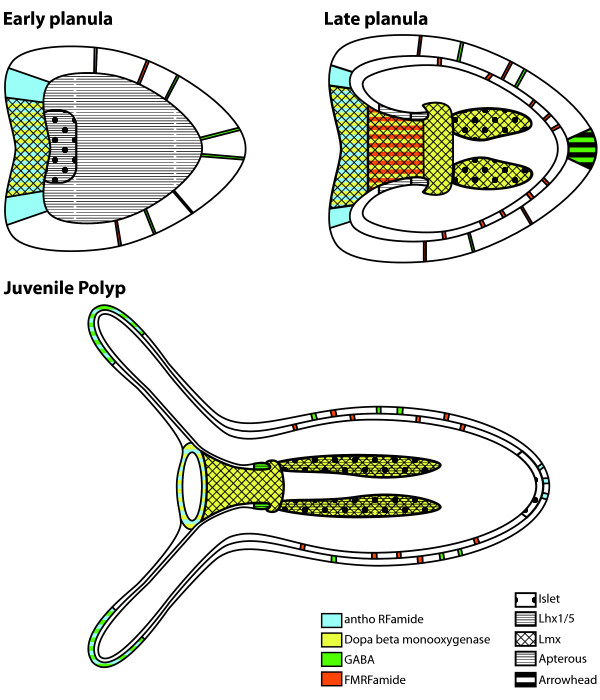
Results: We identified Lhx genes in two cnidarians and a placozoan and report the expression of Lhx genes during embryonic development in Nematostella and the demosponge Amphimedon. Members of the six major LIM homeobox subfamilies are represented in the genomes of the starlet sea anemone, Nematostella vectensis, and the placozoan Trichoplax adhaerens. The hydrozoan cnidarian, Hydra magnipapillata, has retained four of the six Lhx subfamilies, but apparently lost two others. Only three subfamilies are represented in the haplosclerid demosponge Amphimedon queenslandica. A tandem cluster of three Lhx genes of different subfamilies and a gene containing two LIM domains in the genome of T. adhaerens (an animal without any neurons) indicates that Lhx subfamilies were generated by tandem duplication. This tandem cluster in Trichoplax is likely a remnant of the original chromosomal context in which Lhx subfamilies first appeared. Three of the six Trichoplax Lhx genes are expressed in animals in laboratory culture, as are all Lhx genes in Hydra. Expression patterns of Nematostella Lhx genes correlate with neural territories in larval and juvenile polyp stages. In the aneural demosponge, A. queenslandica, the three Lhx genes are expressed widely during development, including in cells that are associated with the larval photosensory ring.
Conclusions: The Lhx family expanded and diversified early in animal evolution, with all six subfamilies already diverged prior to the cnidarian-placozoan-bilaterian last common ancestor. In Nematostella, Lhx gene expression is correlated with neural territories in larval and juvenile polyp stages. This pattern is consistent with a possible role in patterning the Nematostella nervous system. We propose a scenario in which Lhx genes play a homologous role in neural patterning across eumetazoans.
Figures
Similar articles
-
Lim homeobox genes in the Ctenophore Mnemiopsis leidyi: the evolution of neural cell type specification.Evodevo. 2012 Jan 13;3(1):2. doi: 10.1186/2041-9139-3-2. Evodevo. 2012. PMID: 22239757 Free PMC article.
-
A conserved cluster of three PRD-class homeobox genes (homeobrain, rx and orthopedia) in the Cnidaria and Protostomia.Evodevo. 2010 Jul 5;1(1):3. doi: 10.1186/2041-9139-1-3. Evodevo. 2010. PMID: 20849646 Free PMC article.
-
Multiple dicer genes in the early-diverging metazoa.Mol Biol Evol. 2009 Jun;26(6):1333-40. doi: 10.1093/molbev/msp042. Epub 2009 Mar 10. Mol Biol Evol. 2009. PMID: 19276153
-
Cnidarian Zic Genes.Adv Exp Med Biol. 2018;1046:27-39. doi: 10.1007/978-981-10-7311-3_2. Adv Exp Med Biol. 2018. PMID: 29442315 Review.
-
Review: The evolution of peptidergic signaling in Cnidaria and Placozoa, including a comparison with Bilateria.Front Endocrinol (Lausanne). 2022 Sep 23;13:973862. doi: 10.3389/fendo.2022.973862. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36213267 Free PMC article. Review.
Cited by
-
A bipolar role of the transcription factor ERG for cnidarian germ layer formation and apical domain patterning.Dev Biol. 2017 Oct 15;430(2):346-361. doi: 10.1016/j.ydbio.2017.08.015. Epub 2017 Aug 14. Dev Biol. 2017. PMID: 28818668 Free PMC article.
-
Contrasting DNA-binding behaviour by ISL1 and LHX3 underpins differential gene targeting in neuronal cell specification.J Struct Biol X. 2020 Dec 15;5:100043. doi: 10.1016/j.yjsbx.2020.100043. eCollection 2021. J Struct Biol X. 2020. PMID: 33458649 Free PMC article.
-
Dynamics of genomic innovation in the unicellular ancestry of animals.Elife. 2017 Jul 20;6:e26036. doi: 10.7554/eLife.26036. Elife. 2017. PMID: 28726632 Free PMC article.
-
Islet 1 specifies the identity of hypothalamic melanocortin neurons and is critical for normal food intake and adiposity in adulthood.Proc Natl Acad Sci U S A. 2015 Apr 14;112(15):E1861-70. doi: 10.1073/pnas.1500672112. Epub 2015 Mar 30. Proc Natl Acad Sci U S A. 2015. PMID: 25825735 Free PMC article.
-
KEGG orthology-based annotation of the predicted proteome of Acropora digitifera: ZoophyteBase - an open access and searchable database of a coral genome.BMC Genomics. 2013 Jul 26;14:509. doi: 10.1186/1471-2164-14-509. BMC Genomics. 2013. PMID: 23889801 Free PMC article.
References
-
- Greenspan RJ. An introduction to nervous systems. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 2007.
-
- Srivastava M, Begovic E, Chapman J, Putnam NH, Hellsten U, Kawashima T, Kuo A, Mitros T, Salamov A, Carpenter ML, Signorovitch AY, Moreno MA, Kamm K, Grimwood J, Schmutz J, Shapiro H, Grigoriev IV, Buss LW, Schierwater B, Dellaporta SL, Rokhsar DS. The Trichoplax genome and the nature of placozoans. Nature. 2008;454:955–960. doi: 10.1038/nature07191. - DOI - PubMed
-
- Philippe H, Derelle R, Lopez P, Pick K, Borchiellini C, Boury-Esnault N, Vacelet J, Renard E, Houliston E, Queinnec E, Da Silva C, Wincker P, Le Guyader H, Leys S, Jackson DJ, Schreiber F, Erpenbeck D, Morgenstern B, Worheide G, Manuel M. Phylogenomics Revives Traditional Views on Deep Animal Relationships. Curr Biol. 2009. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
