

Coherence potentials: loss-less, all-or-none network events in the cortex
- PMID: 20084093
- PMCID: PMC2795777
- DOI: 10.1371/journal.pbio.1000278
Coherence potentials: loss-less, all-or-none network events in the cortex
Erratum in
- PLoS Biol. 2010;8(1). doi: 10.1371/annotation/668be458-400b-4c19-accf-34fc543340e2
- PLoS Biol. 2010;8(1). doi: 10.1371/annotation/f6193dfe-a90c-4110-b0a0-22d81a87b921
Abstract
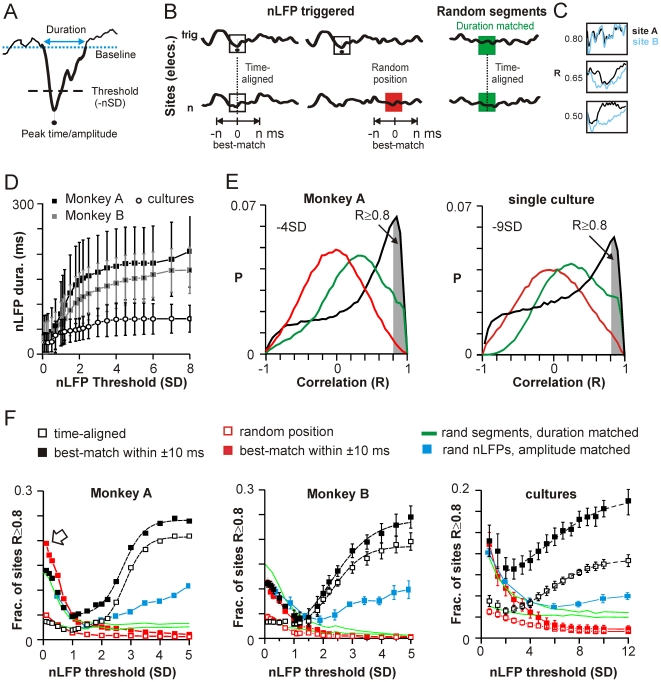
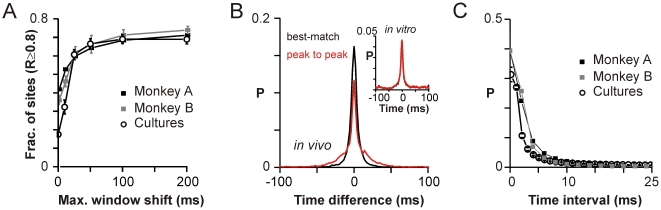
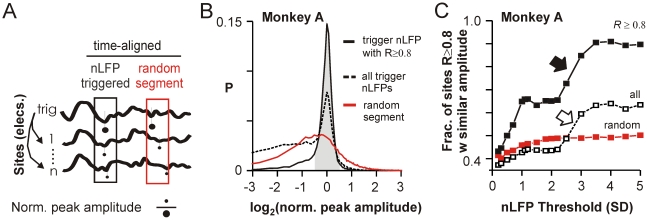
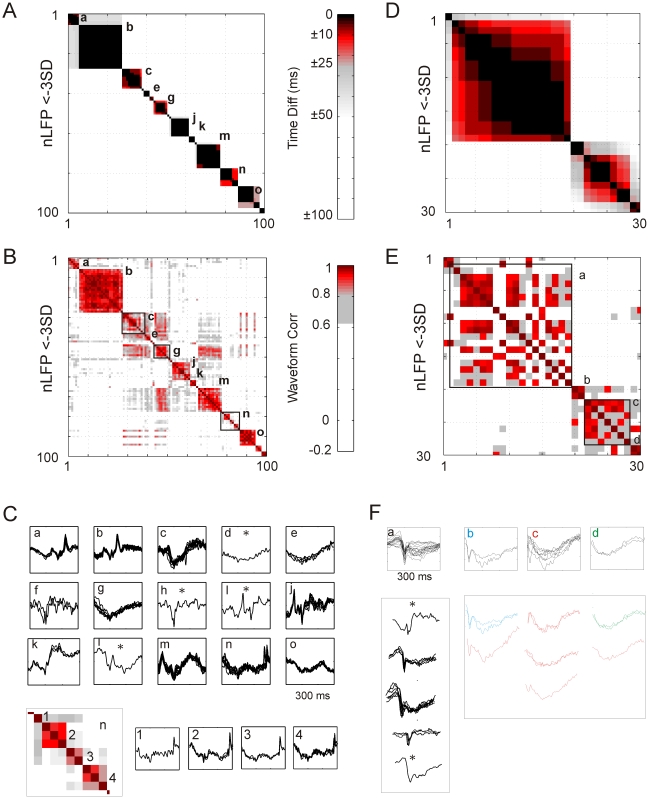
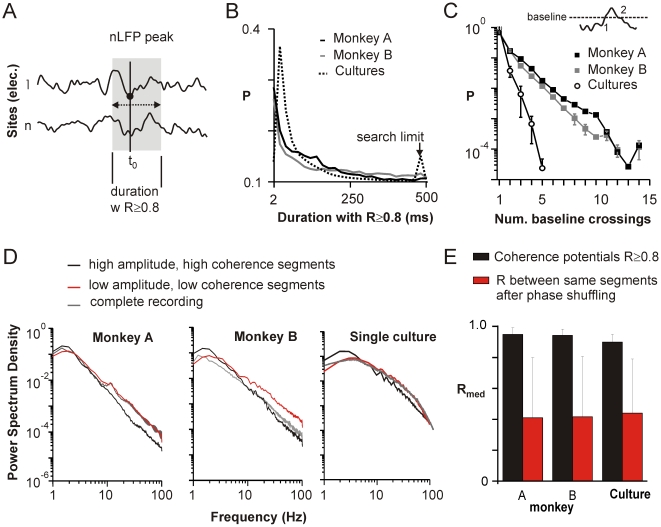
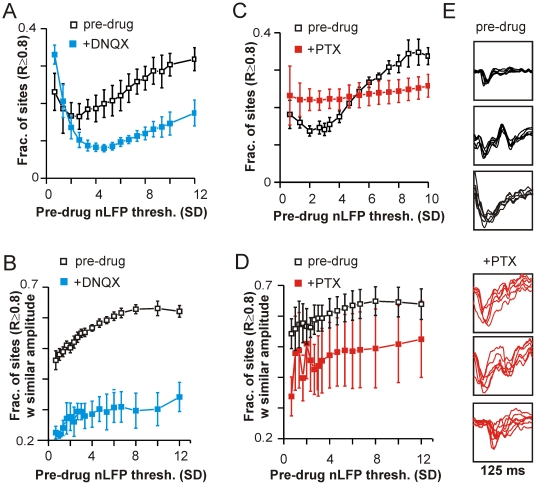
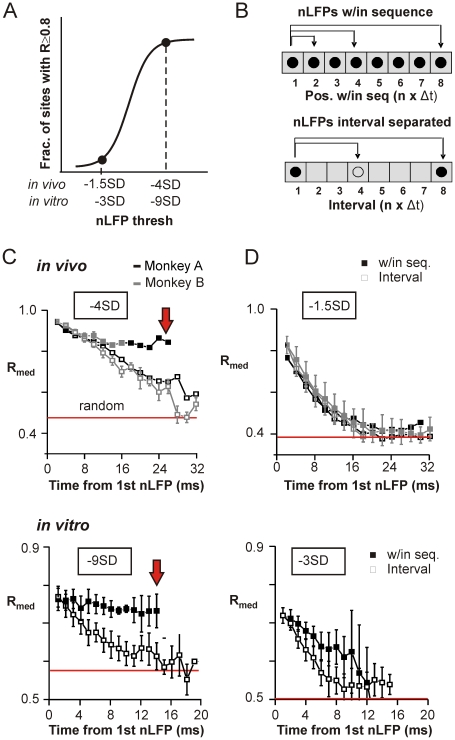
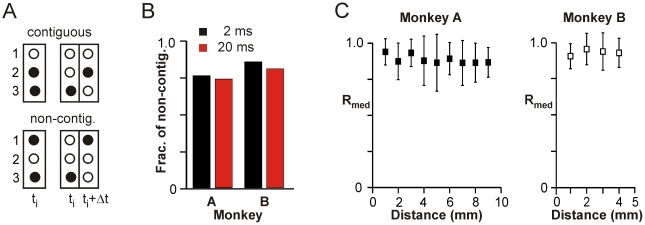
Transient associations among neurons are thought to underlie memory and behavior. However, little is known about how such associations occur or how they can be identified. Here we recorded ongoing local field potential (LFP) activity at multiple sites within the cortex of awake monkeys and organotypic cultures of cortex. We show that when the composite activity of a local neuronal group exceeds a threshold, its activity pattern, as reflected in the LFP, occurs without distortion at other cortex sites via fast synaptic transmission. These large-amplitude LFPs, which we call coherence potentials, extend up to hundreds of milliseconds and mark periods of loss-less spread of temporal and amplitude information much like action potentials at the single-cell level. However, coherence potentials have an additional degree of freedom in the diversity of their waveforms, which provides a high-dimensional parameter for encoding information and allows identification of particular associations. Such nonlinear behavior is analogous to the spread of ideas and behaviors in social networks.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Oscillatory activity in sensorimotor cortex of awake monkeys: synchronization of local field potentials and relation to behavior.J Neurophysiol. 1996 Dec;76(6):3949-67. doi: 10.1152/jn.1996.76.6.3949. J Neurophysiol. 1996. PMID: 8985892
-
Spontaneous cortical activity in awake monkeys composed of neuronal avalanches.Proc Natl Acad Sci U S A. 2009 Sep 15;106(37):15921-6. doi: 10.1073/pnas.0904089106. Epub 2009 Aug 26. Proc Natl Acad Sci U S A. 2009. PMID: 19717463 Free PMC article.
-
Modeling mesoscopic cortical dynamics using a mean-field model of conductance-based networks of adaptive exponential integrate-and-fire neurons.J Comput Neurosci. 2018 Feb;44(1):45-61. doi: 10.1007/s10827-017-0668-2. Epub 2017 Nov 15. J Comput Neurosci. 2018. PMID: 29139050
-
Associative memory in networks of spiking neurons.Neural Netw. 2001 Jul-Sep;14(6-7):825-34. doi: 10.1016/s0893-6080(01)00064-8. Neural Netw. 2001. PMID: 11665774 Review.
-
Dynamic depolarization fields in the cerebral cortex.Trends Neurosci. 2002 Apr;25(4):183-90. doi: 10.1016/s0166-2236(00)02125-1. Trends Neurosci. 2002. PMID: 11998686 Review.
Cited by
-
Critical slowing down governs the transition to neuron spiking.PLoS Comput Biol. 2015 Feb 23;11(2):e1004097. doi: 10.1371/journal.pcbi.1004097. eCollection 2015 Feb. PLoS Comput Biol. 2015. PMID: 25706912 Free PMC article.
-
Scale-invariant neuronal avalanche dynamics and the cut-off in size distributions.PLoS One. 2014 Jun 13;9(6):e99761. doi: 10.1371/journal.pone.0099761. eCollection 2014. PLoS One. 2014. PMID: 24927158 Free PMC article.
-
Tradeoffs and constraints on neural representation in networks of cortical neurons.J Neurosci. 2010 Jul 14;30(28):9588-96. doi: 10.1523/JNEUROSCI.0661-10.2010. J Neurosci. 2010. PMID: 20631187 Free PMC article.
-
Optimal Organization of Functional Connectivity Networks for Segregation and Integration With Large-Scale Critical Dynamics in Human Brains.Front Comput Neurosci. 2021 Mar 31;15:641335. doi: 10.3389/fncom.2021.641335. eCollection 2021. Front Comput Neurosci. 2021. PMID: 33867963 Free PMC article.
-
Fractals in the nervous system: conceptual implications for theoretical neuroscience.Front Physiol. 2010 Jul 6;1:15. doi: 10.3389/fphys.2010.00015. eCollection 2010. Front Physiol. 2010. PMID: 21423358 Free PMC article.
References
-
- Hebb D. O. The organization of behavior. New York: Wiley; 1949.
-
- Bressler S. L, Coppola R, Nakamura R. Episodic multiregional cortical coherence at multiple frequencies during visual task performance. Nature. 1993;366:153–156. - PubMed
-
- Gray C. M, Konig P, Engel A. K, Singer W. Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties. Nature. 1989;338:334–337. - PubMed
-
- Eckhorn R, Bauer R, Jordan W, Brosch M, Kruse W, et al. Coherent oscillations: a mechanism of feature linking in the visual cortex? Multiple electrode and correlation analyses in the cat. Biol Cybern. 1988;60:121–130. - PubMed
-
- Aertsen A. M, Gerstein G. L, Habib M. K, Palm G. Dynamics of neuronal firing correlation: modulation of “effective connectivity”. J Neurophysiol. 1989;61:900–917. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
