

Control of Alzheimer's amyloid beta toxicity by the high molecular weight immunophilin FKBP52 and copper homeostasis in Drosophila
- PMID: 20084280
- PMCID: PMC2801609
- DOI: 10.1371/journal.pone.0008626
Control of Alzheimer's amyloid beta toxicity by the high molecular weight immunophilin FKBP52 and copper homeostasis in Drosophila
Abstract
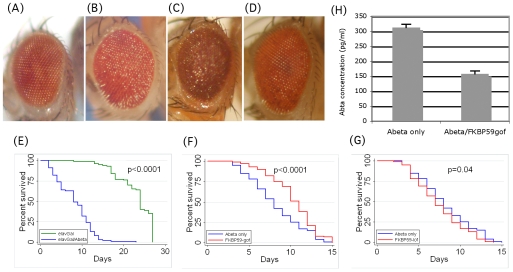
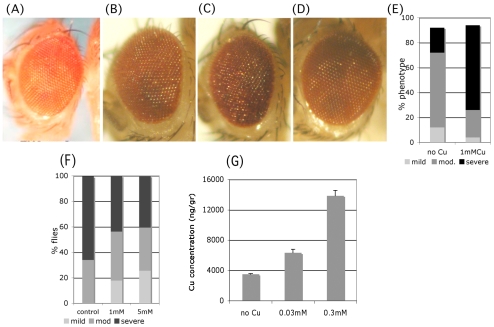
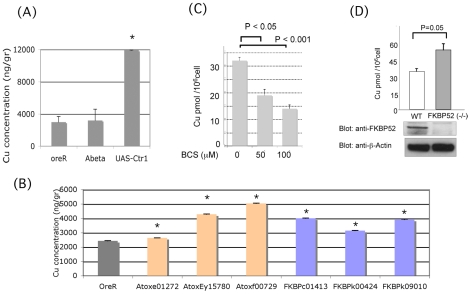
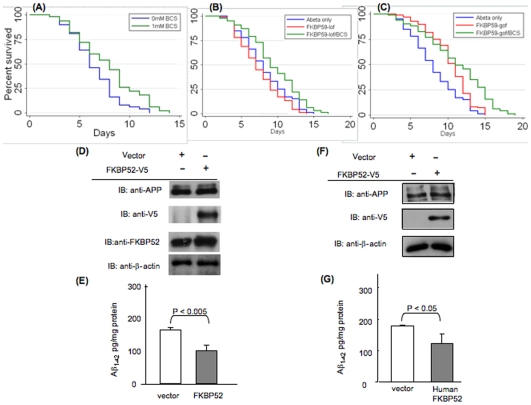
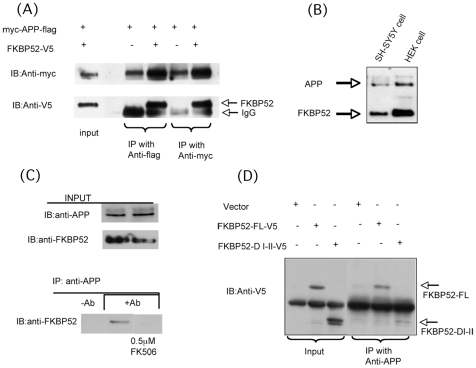
FK506 binding proteins (FKBPs), also called immunophilins, are prolyl-isomerases (PPIases) that participate in a wide variety of cellular functions including hormone signaling and protein folding. Recent studies indicate that proteins that contain PPIase activity can also alter the processing of Alzheimer's Amyloid Precursor Protein (APP). Originally identified in hematopoietic cells, FKBP52 is much more abundantly expressed in neurons, including the hippocampus, frontal cortex, and basal ganglia. Given the fact that the high molecular weight immunophilin FKBP52 is highly expressed in CNS regions susceptible to Alzheimer's, we investigated its role in Abeta toxicity. Towards this goal, we generated Abeta transgenic Drosophila that harbor gain of function or loss of function mutations of FKBP52. FKBP52 overexpression reduced the toxicity of Abeta and increased lifespan in Abeta flies, whereas loss of function of FKBP52 exacerbated these Abeta phenotypes. Interestingly, the Abeta pathology was enhanced by mutations in the copper transporters Atox1, which interacts with FKBP52, and Ctr1A and was suppressed in FKBP52 mutant flies raised on a copper chelator diet. Using mammalian cultures, we show that FKBP52 (-/-) cells have increased intracellular copper and higher levels of Abeta. This effect is reversed by reconstitution of FKBP52. Finally, we also found that FKBP52 formed stable complexes with APP through its FK506 interacting domain. Taken together, these studies identify a novel role for FKBP52 in modulating toxicity of Abeta peptides.
Conflict of interest statement
Figures
Similar articles
-
A novel role for the immunophilin FKBP52 in copper transport.J Biol Chem. 2004 Jul 2;279(27):27845-8. doi: 10.1074/jbc.C400118200. Epub 2004 May 7. J Biol Chem. 2004. PMID: 15133031
-
A novel role for the immunophilin FKBP52 in motor coordination.Behav Brain Res. 2016 Oct 15;313:97-110. doi: 10.1016/j.bbr.2016.07.015. Epub 2016 Jul 11. Behav Brain Res. 2016. PMID: 27418439
-
FKBP immunophilins and Alzheimer's disease: a chaperoned affair.J Biosci. 2011 Aug;36(3):493-8. doi: 10.1007/s12038-011-9080-7. J Biosci. 2011. PMID: 21799260 Review.
-
Proof that the high molecular weight immunophilin FKBP52 mediates the in vivo neuroregenerative effect of the macrolide FK506.Biochem Pharmacol. 2020 Dec;182:114204. doi: 10.1016/j.bcp.2020.114204. Epub 2020 Aug 20. Biochem Pharmacol. 2020. PMID: 32828804
-
Comparative analysis of FKBP family protein: evaluation, structure, and function in mammals and Drosophila melanogaster.BMC Dev Biol. 2018 Mar 27;18(1):7. doi: 10.1186/s12861-018-0167-3. BMC Dev Biol. 2018. PMID: 29587629 Free PMC article. Review.
Cited by
-
HSP90AB1: Helping the good and the bad.Gene. 2016 Jan 10;575(2 Pt 1):171-86. doi: 10.1016/j.gene.2015.08.063. Epub 2015 Sep 7. Gene. 2016. PMID: 26358502 Free PMC article. Review.
-
Copper Redox Cycling Inhibits Aβ Fibre Formation and Promotes Fibre Fragmentation, while Generating a Dityrosine Aβ Dimer.Sci Rep. 2018 Nov 1;8(1):16190. doi: 10.1038/s41598-018-33935-5. Sci Rep. 2018. PMID: 30385800 Free PMC article.
-
Copper and Zinc Homeostasis: Lessons from Drosophila melanogaster.Front Genet. 2017 Dec 21;8:223. doi: 10.3389/fgene.2017.00223. eCollection 2017. Front Genet. 2017. PMID: 29312444 Free PMC article. Review.
-
Conserved metallomics in two insect families evolving separately for a hundred million years.Biometals. 2014 Dec;27(6):1323-35. doi: 10.1007/s10534-014-9793-9. Epub 2014 Oct 9. Biometals. 2014. PMID: 25298233 Free PMC article.
-
Truncated Amyloid-β(11-40/42) from Alzheimer Disease Binds Cu2+ with a Femtomolar Affinity and Influences Fiber Assembly.J Biol Chem. 2015 Nov 13;290(46):27791-802. doi: 10.1074/jbc.M115.684084. Epub 2015 Sep 25. J Biol Chem. 2015. PMID: 26408196 Free PMC article.
References
-
- Tai PK, Maeda Y, Nakao K, Wakim NG, Duhring JL, et al. A 59-killodalton protein associated with progestin, estrogen, androgen, and glucocorticoid receptor. Biochemistry. 1986;25:5269–5275. - PubMed
-
- Steiner JP, Dawson TM, Fotuhi M, Glatt CE, Snowman AM, et al. High brain densities of the immunophilin FKBP colocalized with calcineurin. Nature. 1992;358(6387):584–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
