

A quantitative assessment of torque-transducer models for magnetoreception
- PMID: 20086054
- PMCID: PMC2843997
- DOI: 10.1098/rsif.2009.0435.focus
A quantitative assessment of torque-transducer models for magnetoreception
Abstract
Although ferrimagnetic material appears suitable as a basis of magnetic field perception in animals, it is not known by which mechanism magnetic particles may transduce the magnetic field into a nerve signal. Provided that magnetic particles have remanence or anisotropic magnetic susceptibility, an external magnetic field will exert a torque and may physically twist them. Several models of such biological magnetic-torque transducers on the basis of magnetite have been proposed in the literature. We analyse from first principles the conditions under which they are viable. Models based on biogenic single-domain magnetite prove both effective and efficient, irrespective of whether the magnetic structure is coupled to mechanosensitive ion channels or to an indirect transduction pathway that exploits the strayfield produced by the magnetic structure at different field orientations. On the other hand, torque-detector models that are based on magnetic multi-domain particles in the vestibular organs turn out to be ineffective. Also, we provide a generic classification scheme of torque transducers in terms of axial or polar output, within which we discuss the results from behavioural experiments conducted under altered field conditions or with pulsed fields. We find that the common assertion that a magnetoreceptor based on single-domain magnetite could not form the basis for an inclination compass does not always hold.
Figures
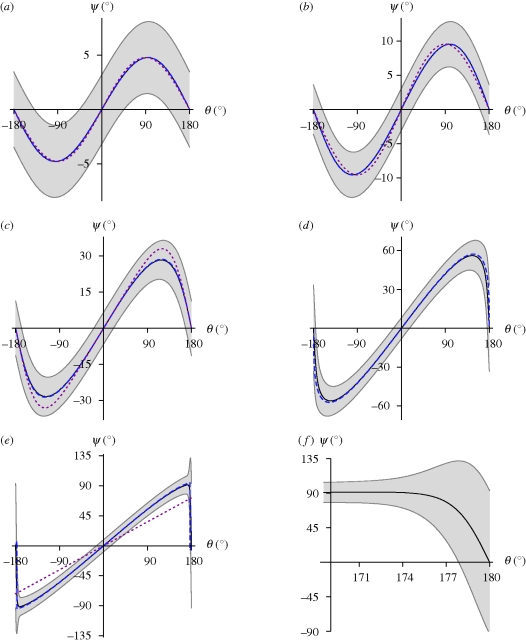
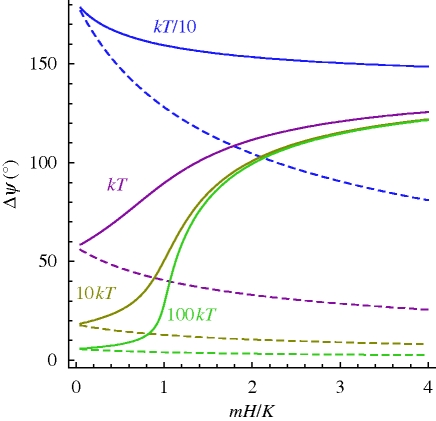
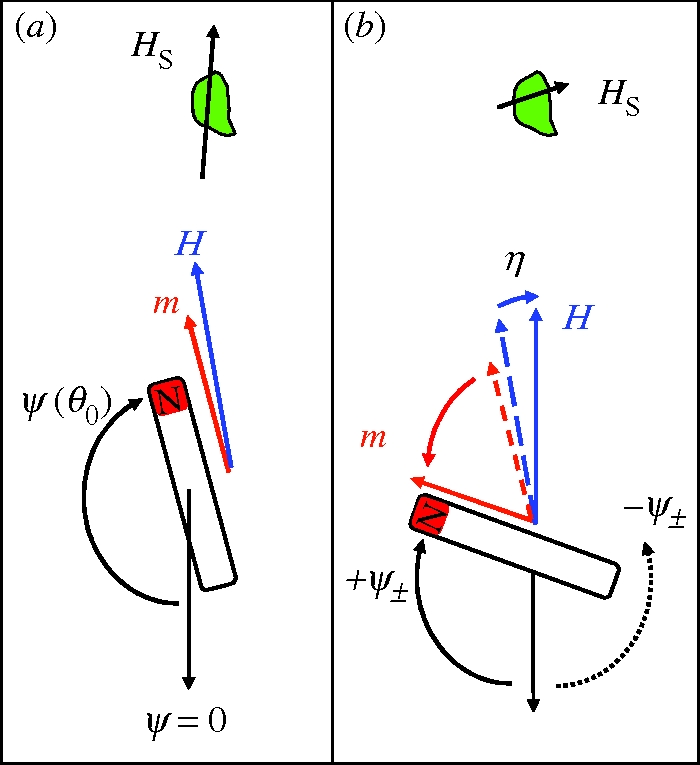
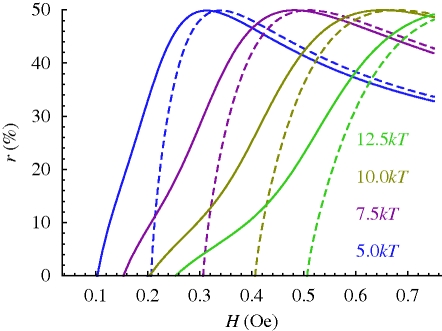
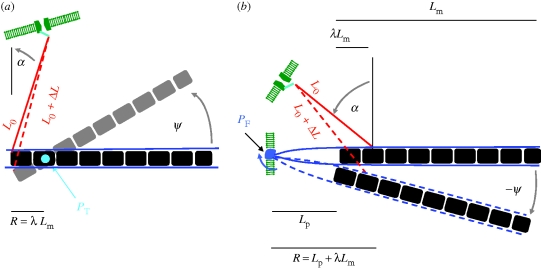
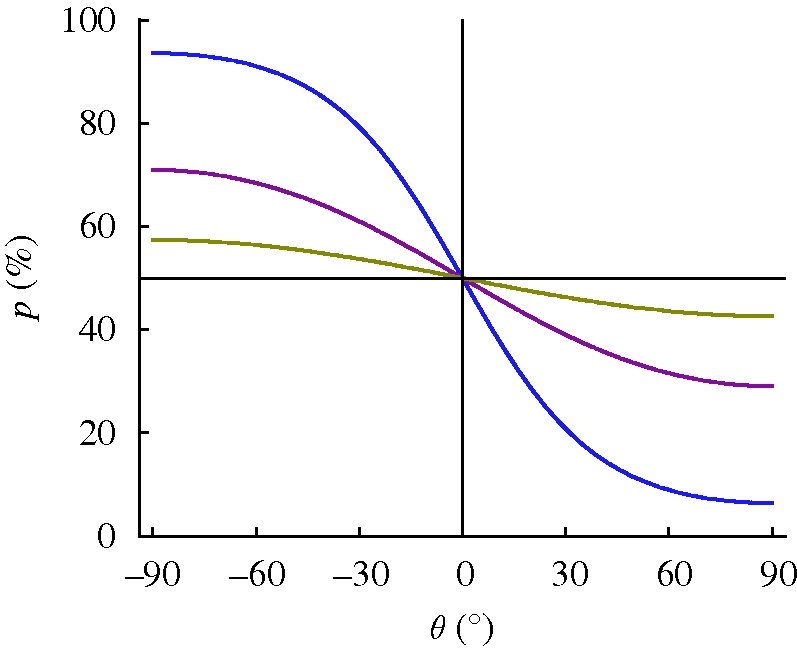
References
-
- Bazylinski D. A., Schlezinger D. R., Howes B. H., Frankel R. B., Epstein S. S. 2000. Occurrence and distribution of diverse populations of magnetic protists in a chemically stratified coastal salt pond. Chem. Geol. 169, 319–328. ( 10.1016/S0009-2541(00)00211-4) - DOI
-
- Beason R. C., Wiltschko R., Wiltschko W. 1997. Pigeon homing: effects of magnetic pulses on initial orientation. Auk 114, 405–415.
-
- Boulbitch A. A. 1998. Deflection of a cell membrane under application of a local force. Phys. Rev. E 57, 2123–2128. ( 10.1103/PhysRevE.57.2123) - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources