

Control of cell survival and proliferation by mammalian eukaryotic initiation factor 4B
- PMID: 20086100
- PMCID: PMC2832492
- DOI: 10.1128/MCB.01218-09
Control of cell survival and proliferation by mammalian eukaryotic initiation factor 4B
Abstract
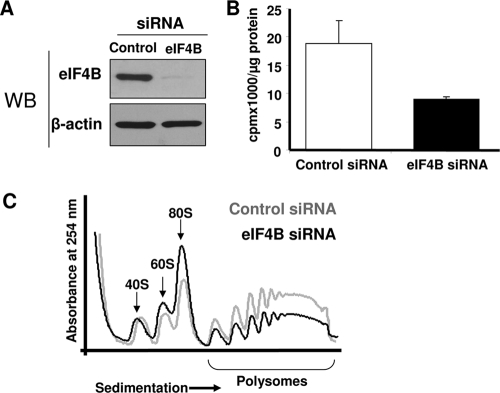
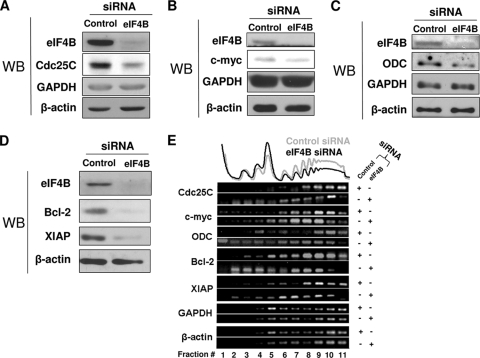
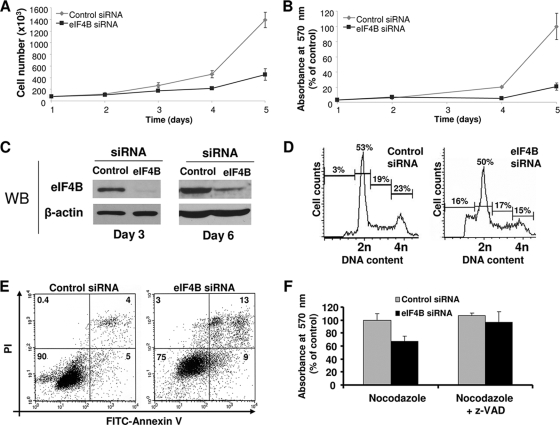
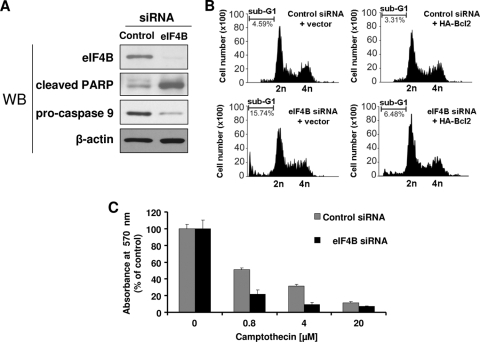
Translation initiation plays an important role in cell growth, proliferation, and survival. The translation initiation factor eIF4B (eukaryotic initiation factor 4B) stimulates the RNA helicase activity of eIF4A in unwinding secondary structures in the 5' untranslated region (5'UTR) of the mRNA in vitro. Here, we studied the effects of eIF4B depletion in cells using RNA interference (RNAi). In agreement with the role of eIF4B in translation initiation, its depletion resulted in inhibition of this step. Selective reduction of translation was observed for mRNAs harboring strong to moderate secondary structures in their 5'UTRs. These mRNAs encode proteins, which function in cell proliferation (Cdc25C, c-myc, and ODC [ornithine decarboxylase]) and survival (Bcl-2 and XIAP [X-linked inhibitor of apoptosis]). Furthermore, eIF4B silencing led to decreased proliferation rates, promoted caspase-dependent apoptosis, and further sensitized cells to camptothecin-induced cell death. These results demonstrate that eIF4B is required for cell proliferation and survival by regulating the translation of proliferative and prosurvival mRNAs.
Figures
Similar articles
-
eIF4B stimulates translation of long mRNAs with structured 5' UTRs and low closed-loop potential but weak dependence on eIF4G.Proc Natl Acad Sci U S A. 2016 Sep 20;113(38):10464-72. doi: 10.1073/pnas.1612398113. Epub 2016 Sep 6. Proc Natl Acad Sci U S A. 2016. PMID: 27601676 Free PMC article.
-
eIF4B stimulates eIF4A ATPase and unwinding activities by direct interaction through its 7-repeats region.RNA Biol. 2017 Jan 2;14(1):113-123. doi: 10.1080/15476286.2016.1259782. Epub 2016 Nov 18. RNA Biol. 2017. PMID: 27858515 Free PMC article.
-
Eukaryotic initiation factor 4B is a multi-functional RNA binding protein that regulates histone mRNAs.Nucleic Acids Res. 2024 Oct 28;52(19):12039-12054. doi: 10.1093/nar/gkae767. Nucleic Acids Res. 2024. PMID: 39225047 Free PMC article.
-
mRNA helicases: the tacticians of translational control.Nat Rev Mol Cell Biol. 2011 Apr;12(4):235-45. doi: 10.1038/nrm3083. Nat Rev Mol Cell Biol. 2011. PMID: 21427765 Review.
-
eIF4F: a retrospective.J Biol Chem. 2015 Oct 2;290(40):24091-9. doi: 10.1074/jbc.R115.675280. Epub 2015 Aug 31. J Biol Chem. 2015. PMID: 26324716 Free PMC article. Review.
Cited by
-
Translating from cancer to the brain: regulation of protein synthesis by eIF4F.Learn Mem. 2019 Aug 15;26(9):332-342. doi: 10.1101/lm.050047.119. Print 2019 Sep. Learn Mem. 2019. PMID: 31416906 Free PMC article. Review.
-
Inhibition of melanocortin 1 receptor slows melanoma growth, reduces tumor heterogeneity and increases survival.Oncotarget. 2016 May 3;7(18):26331-45. doi: 10.18632/oncotarget.8372. Oncotarget. 2016. PMID: 27028866 Free PMC article.
-
Mitotic MELK-eIF4B signaling controls protein synthesis and tumor cell survival.Proc Natl Acad Sci U S A. 2016 Aug 30;113(35):9810-5. doi: 10.1073/pnas.1606862113. Epub 2016 Aug 15. Proc Natl Acad Sci U S A. 2016. PMID: 27528663 Free PMC article.
-
The Role of Translational Regulation in Survival after Radiation Damage; an Opportunity for Proteomics Analysis.Proteomes. 2014 Jun;2(2):272-290. doi: 10.3390/proteomes2020272. Proteomes. 2014. PMID: 26269784 Free PMC article.
-
Differential Proteome Analysis of Human Neuroblastoma Xenograft Primary Tumors and Matched Spontaneous Distant Metastases.Sci Rep. 2018 Sep 18;8(1):13986. doi: 10.1038/s41598-018-32236-1. Sci Rep. 2018. PMID: 30228356 Free PMC article.
References
-
- Bedikian, A. Y., M. Millward, H. Pehamberger, R. Conry, M. Gore, U. Trefzer, A. C. Pavlick, R. DeConti, E. M. Hersh, P. Hersey, J. M. Kirkwood, and F. G. Haluska. 2006. Bcl-2 antisense (oblimersen sodium) plus dacarbazine in patients with advanced melanoma: the Oblimersen Melanoma Study Group. J. Clin. Oncol. 24:4738-4745. - PubMed
-
- Benne, R., and J. W. Hershey. 1978. The mechanism of action of protein synthesis initiation factors from rabbit reticulocytes. J. Biol. Chem. 253:3078-3087. - PubMed
-
- Benz, E. J., Jr., D. G. Nathan, R. K. Amaravadi, and N. N. Danial. 2007. Targeting the cell death-survival equation. Clin. Cancer Res. 13:7250-7253. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous