

Rethinking the Warburg effect with Myc micromanaging glutamine metabolism
- PMID: 20086171
- PMCID: PMC2818441
- DOI: 10.1158/0008-5472.CAN-09-3556
Rethinking the Warburg effect with Myc micromanaging glutamine metabolism
Abstract
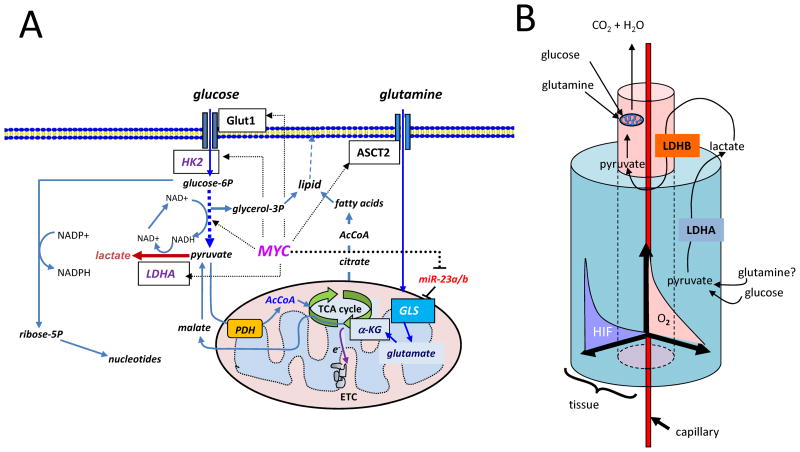
The MYC oncogene, which is frequently deregulated in human cancers, encodes a master transcription factor c-Myc (herein termed Myc) that integrates cell proliferation with metabolism through its regulation of thousands of genes including microRNAs (miRNA). In addition to its known function in regulating the cell cycle and glucose metabolism, recent studies document a role for Myc in stimulating glutamine catabolism, in part through the repression of miRNAs miR-23a and miR-23b. These observations suggest an additional level of complexity in tumor metabolism, which includes the commensal metabolic relationship between hypoxic and nonhypoxic regions of tumors as well as the surrounding stroma. Thus, a reevaluation of cancer metabolism considering glutamine catabolism with a better understanding of the tumor histological complexity is needed before cancer metabolism can be effectively targeted in therapy.
Figures
References
-
- Kroemer G, Pouyssegur J. Tumor cell metabolism: cancer’s Achilles’ heel. Cancer Cell. 2008;13:472–482. - PubMed
-
- Kim JW, Dang CV. Cancer’s molecular sweet tooth and the Warburg effect. Cancer Res. 2006;66:8927–8930. - PubMed
-
- Parsons DW, Jones S, Zhang X, Lin JC, Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, Olivi A, McLendon R, Rasheed BA, Keir S, Nikolskaya T, Nikolsky Y, Busam DA, Tekleab H, Diaz LA, Jr, Hartigan J, Smith DR, Strausberg RL, Marie SK, Shinjo SM, Yan H, Riggins GJ, Bigner DD, Karchin R, Papadopoulos N, Parmigiani G, Vogelstein B, Velculescu VE, Kinzler KW. An integrated genomic analysis of human glioblastoma multiforme. Science. 2008;321:1807–1812. - PMC - PubMed
-
- Flier JS, Mueckler MM, Usher P, Lodish HF. Elevated levels of glucose transport and transporter messenger RNA are induced by ras or src oncogenes. Science. 1987;235:1492–1495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
