

Distinct signal codes generate dendritic cell functional plasticity
- PMID: 20086239
- PMCID: PMC3325779
- DOI: 10.1126/scisignal.2000567
Distinct signal codes generate dendritic cell functional plasticity
Abstract
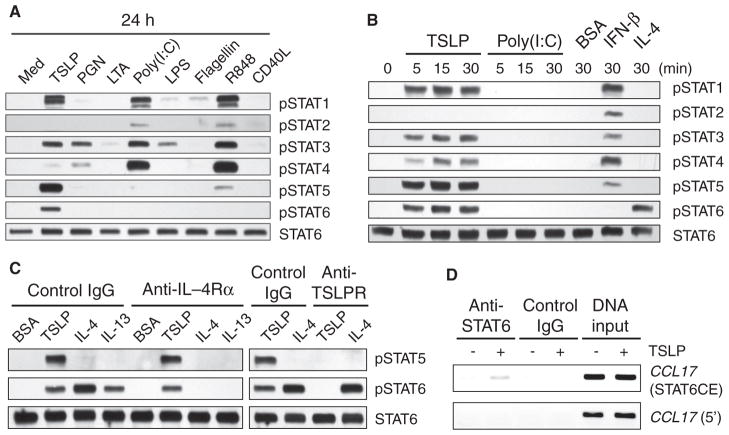
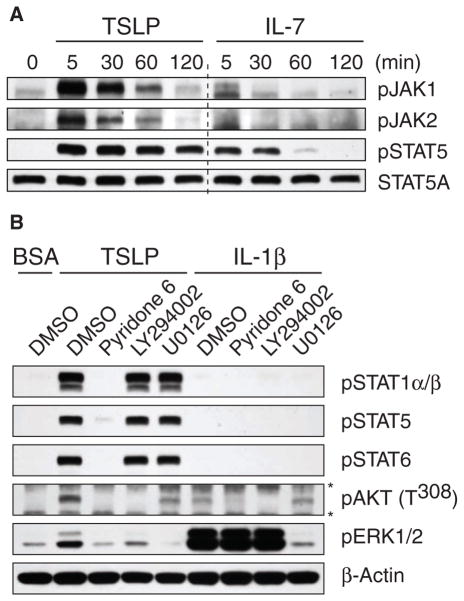
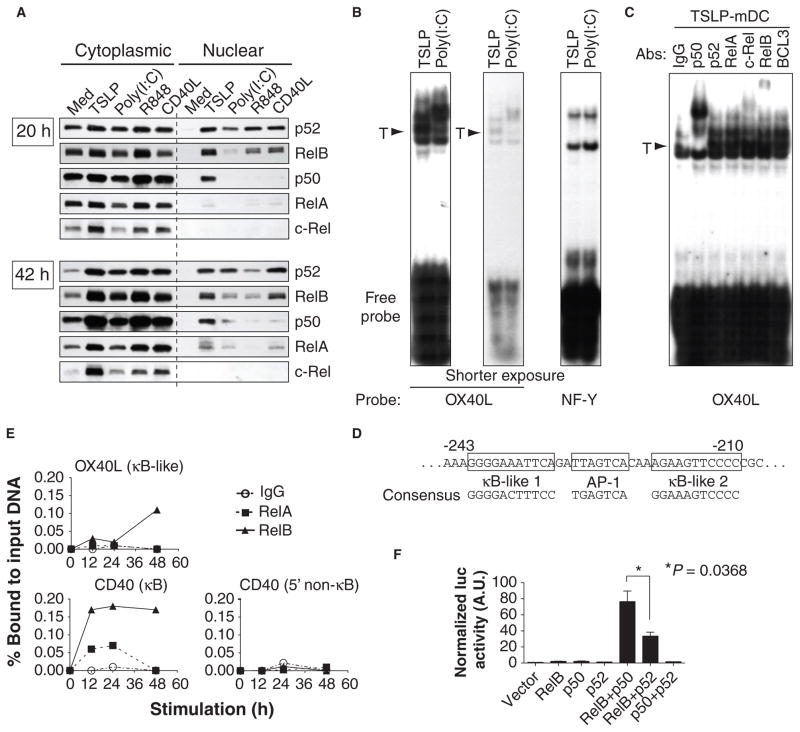
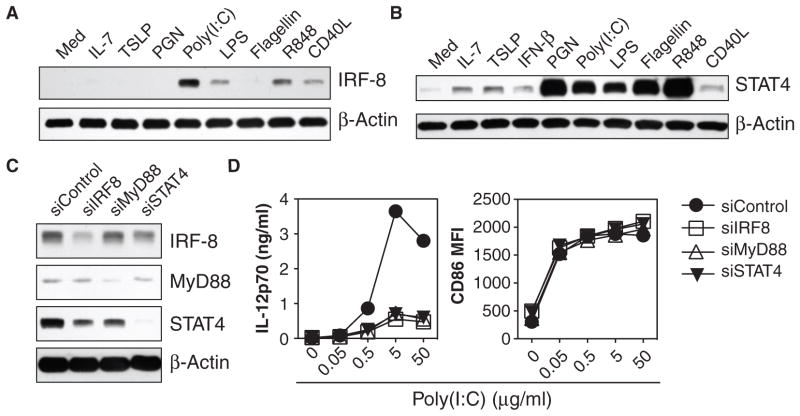
Our adaptive immune system induces distinct responses to different pathogens because of the functional plasticity of dendritic cells (DCs); however, how DCs program unique responses remains unclear. Here, we found that the cytokine thymic stromal lymphopoietin (TSLP) potently transduced a unique T helper type 2 (T(H)2)-inducing compound signal in DCs. Whereas activation of nuclear factor kappaB (predominantly p50) drove DCs to produce OX40L to induce T(H)2 differentiation, the activation of signal transducer and activator of transcription 6 (STAT6) triggered DCs to secrete chemokines necessary for the recruitment of T(H)2 cells. In addition, TSLP signaling limited the activation of STAT4 and interferon regulatory factor 8 (IRF-8), which are essential factors for the production of the T(H)1-polarizing cytokine interleukin-12 (IL-12). By contrast, Toll-like receptor ligands and CD40 ligand did not activate STAT6 in myeloid DCs, but instead increased the abundance of STAT4 and IRF-8 to induce T(H)1 responses through the production of IL-12. Therefore, we propose that the functional plasticity of DCs relies on elaborate signal codes that are generated by different stimuli.
Figures
References
-
- Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
-
- Macatonia SE, Hosken NA, Litton M, Vieira P, Hsieh CS, Culpepper JA, Wysocka M, Trinchieri G, Murphy KM, O’Garra A. Dendritic cells produce IL-12 and direct the development of Th1 cells from naive CD4+ T cells. J Immunol. 1995;154:5071–5079. - PubMed
-
- Kaliński P, Hilkens CM, Wierenga EA, Kapsenberg ML. T-cell priming by type-1 and type-2 polarized dendritic cells: The concept of a third signal. Immunol Today. 1999;20:561–567. - PubMed
-
- Moser M, Murphy KM. Dendritic cell regulation of TH1-TH2 development. Nat Immunol. 2000;1:199–205. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
